Congenital malformations of the heart are frequent and are found in nearly 1% of all newborns. The complexity of cardiac development is also demonstrated by the fact that many knockout mutations have a cardiac phenotype. In this project we are trying to characterize signalling molecules, transcription factors and other genes with cardiac-restricted expression pattern, that are involved in early organogenesis of the vertebrate heart. We are using both the chicken and mouse embryo as model systems. Currently we are working on two problems:
1. Molecular determinants of left-right development The heart is the first organ to be formed in the body. It is also the first to display L-R asymmetry. Soon after formation of the linear primary heart tube, the heart bends, or loops to the right (Fig. 1).
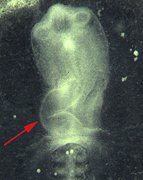 Fig. 1Manipulation of heart looping. Chick embryos are shown after overnight incubation in New culture. Embryos were implanted at stage 4 with a pellet of fibroblasts infected with a Lefty2 virus. After the manipulation embryos displayed either normal looping of the heart to the right (left), unlooped, or symmetric hearts (middle) or showed inversed leftward looping (right). Arrows point to the direction of heart looping.
Fig. 1Manipulation of heart looping. Chick embryos are shown after overnight incubation in New culture. Embryos were implanted at stage 4 with a pellet of fibroblasts infected with a Lefty2 virus. After the manipulation embryos displayed either normal looping of the heart to the right (left), unlooped, or symmetric hearts (middle) or showed inversed leftward looping (right). Arrows point to the direction of heart looping.
A signalling cascade has been identified in vertebrate embryos (Fig. 2A) which mediates left-right (L-R) asymmetry from the region of the organizer (Hensen's node in the chick) to the lateral plate mesoderm (LPM). Most organs displaying L-R asymmetry including the
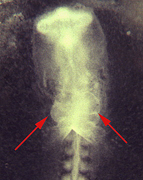 Fig. 2Molecular pathways controlling left-right (L-R) asymmetry and heart field formation. A) Generation of L-R asymmetry by a signalling cascade involving Shh and several TGF? superfamily members, such as nodal, Lefty1 and Lefty2. Ultimately two homeobox genes are induced in the lateral plate mesoderm, Pitx2 and NKX3.2, which are probably involved in asymmetric organ morphogenesis. B) Cardiac mesoderm formation is induced by BMP2 signalling and simultaneous inhibition of Wnt8 by the soluble frizzled receptor crescent. Cardiac mesoderm formation is characterized by the induction of the homeobox gene NKX2.5 and the zinc finger transcription factor GATA4.
Fig. 2Molecular pathways controlling left-right (L-R) asymmetry and heart field formation. A) Generation of L-R asymmetry by a signalling cascade involving Shh and several TGF? superfamily members, such as nodal, Lefty1 and Lefty2. Ultimately two homeobox genes are induced in the lateral plate mesoderm, Pitx2 and NKX3.2, which are probably involved in asymmetric organ morphogenesis. B) Cardiac mesoderm formation is induced by BMP2 signalling and simultaneous inhibition of Wnt8 by the soluble frizzled receptor crescent. Cardiac mesoderm formation is characterized by the induction of the homeobox gene NKX2.5 and the zinc finger transcription factor GATA4.
heart are derivatives of the LPM. Cardiac mesoderm is specified around stages 4-5 and specification includes BMP2 signalling from the adjacent mesendoderm and probably localized inhibition of Wnt signalling by solubilized frizzled receptors (Fig. 2B). An unresolved issue is, how the heart receives it laterality instructions. Heart specification occurs at stages 4-5 while the laterality cascade does not reach the LPM before stages 6-7. Using various agonists and inhibitors we are trying to understand how L-R signalling and cardiac specification are integrated by cardiac mesoderm. Currently we are focusing on the signalling molecul Cryptic which is specifically expressed in the precardiac mesoderm. Overexpression of Cryptic affects heart looping resulting in symmetric hearts or hearts with reversed looping. Looping is a decisive process in cardiac organogenesis. After looping has been initiated chamber-specific gene expression becomes first apparent. Genes controlling heart looping are currently unknown and are the focus of a screen using subtractive hybridization.
2. Isolation of cardiac-restricted genes using subtractive hybridization In order to isolate novel cDNAs specifically expressed in the developing heart a subtractive hybridization approach was utilized. One of the isolated clones was specifically expressed in the chicken heart. Extensive database searches revealed the existence of a gene family consisting of three members in mouse and man, while only two different cDNAs were found in chicken. Northern blot analysis revealed expression in adult skeletal muscle and heart, while minor levels were found in lung and pregnant uterus. Based on the preferential expression in striated muscle this gene family was termed 'Popeye' (POP) genes. POP genes have three transmembrane domains, which are conserved in all family members. Interestingly POP genes displayed several interesting features in their expression pattern. POP3 in chicken and Pop1 in mouse were preferentially expressed in the atrium during the first days of cardiac development. Coincident with epicardial covering of the ventricles both mouse Pop1 and chicken POP3 were expressed in the ventricle confined to the subepicardial compact layer (Fig. 3). Another family member in each species, POP1 in chicken and POP2 in mouse were expressed in the whole heart with the exception of the outflow tract. In human heart POP genes were differentially expressed in the various cardiac chambers. Both human POP2 and POP3 were expressed at high levels in left ventricle, at intermediate levels in the right ventricle and the least in atria. Further work is required to define the function of this novel gene family in cardiac development.
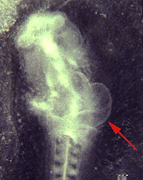 Fig. 3 Expression of Pop1 in the subepicardial compact layer. A) Both atria and ventricle display Pop1 expression, however the outflow tract does not express Pop1. B) In transverse sections Pop1 is found confined to the subepicardial compact layer. C) LacZ staining of a newborn heart of a chimera harboring a lacZ knock in allel of Pop1. D) Transverse section through the ventricle displaying expression in the newborn heart confined to the subepicardial layer.
Fig. 3 Expression of Pop1 in the subepicardial compact layer. A) Both atria and ventricle display Pop1 expression, however the outflow tract does not express Pop1. B) In transverse sections Pop1 is found confined to the subepicardial compact layer. C) LacZ staining of a newborn heart of a chimera harboring a lacZ knock in allel of Pop1. D) Transverse section through the ventricle displaying expression in the newborn heart confined to the subepicardial layer.
People involved in the work - Birgit AndrÈe, Ph.D. student (Isolation and characterisation of the Popeye family,. Generation of a Pop1 knockout mouse). - Thomas Schlange, Ph.D. student (Functional characterization of Cryptic in the early chicken embryo. Integration of L-R signalling and heart field formation) - Bianca Macke, Diploma student (Isolation of looping genes by subtractive hybridization) - Ingo Schnipkoweit, Diploma student (Characterization of the Pop2 locus) - Kerstin Zander, technician (whole mount in situ hybridization, sequencing, large scale gene expression screen)
-
Recent publications - Brand T., Andrée, B., Schneider, A., Buchberger, A., Arnold, H.-H. (1997). Chicken NKx2-8, a novel homeobox gene expressed during early heart and foregut development. Mechanisms of Development 64, 53-59.
-
Andrée, B., Duprez, D., Vorbusch, B., Arnold, H.-H., Brand, T. (1998). BMP-2 induces ectopic expression of cardiac lineage markers and interferes with somite formation in chicken embryos. Mechanisms of Development 70, 119-131.
-
Schneider, A., Mijalski, T., Schlange, T., Dai, W., Overbeek, P., Arnold, H.-H., Brand, T. (1999). The homeobox gene NKX3.2 is a target of left-right signalling and is expressed on opposite sides in chicke and mouse embryos. Current Biology 9, 911-914.
-
Brand, T. Arnold H.-H., Andrée, B. (1999) Entwicklungsgenetische Störungen in der Kardiologie: Zusammenwirken von Grundlagenforschung und Klinik. Medizinische Genetik 11, 250-256.
-
Schlange, T., Andrée, B., Arnold, H.-H., Brand, T. (2000). Distinct requirement of BMP2 for myocardial marker gene expression in the forming heart field in chicken embryos. Mechanisms of Development (in the press).
-
AndrÈe, B., Hillemann, T., Kessler-Icekson, G., Schlesinger, H., Vidne, B.A., Schmitt-John, T., Jockusch, H., Arnold, H.-H., Brand,T. (2000). Isolation and characterization of the novel Popeye gene family expre ssed in skeletal muscle and heart. (submitted).