
Home > Phylum selection > General information
Home How to cite this site Terms & conditions Disclaimer Contact Site tutorial / Help Links |
Monophyly - Paraphyly - Polyphyly An introduction |
The figure below shows a collection of
objects of different shapes and colors. We can make groups of objects according to criteria that we choose and then give names to the groups. For example, we can group together all objects with the same shape:  Choosing the shape as the criterion for group formation leads to four groups of objects. We could also choose the color as the criterion to form groups from the initial set of objects: 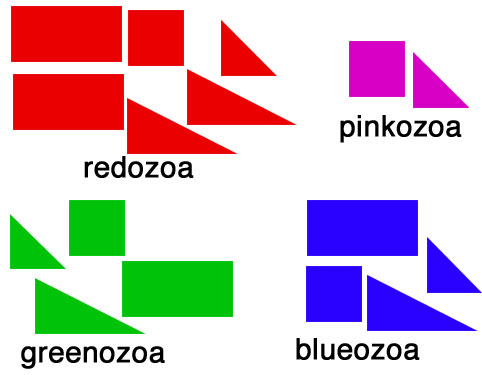 This criterion also leads to four groups of objects, but these are different from the groups formed before. We have now two different systems for grouping the objects. The criterion for system 1 is the shape of the objects, and the criterion for system 2 is the color of the objects. Neither system is better or worse than the other, and none is "the correct" system, since the criteria (shape, color) were chosen arbitrarily. Systems like these are called "artificial" or "non-natural". However, in some cases there can be a preexisting system that links the objects. This system is not man-made and is therefore called "natural". The objects might be linked, because the descended from each other: 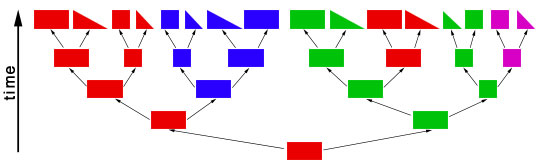 In this scenario the objects have evolved in several steps from a red rectangle. In the first generation, the red rectangle produced two offspring: one of them retained the original shape and color, but the second offspring evolved a new color, green. Evolutionary changes of shape and color also occurred in the next generations, leading to the set of objects that we already know from our previous considerations. The objects are now linked by descent and, of course, we now want to make groups that join together related objects, in order to obtain "groups of relatives", "families" or "natural groups". Two groups in this tree already appeared in our previous systems: all pink objects derive from a common ancestor and all blue objects derive from a common ancestor: 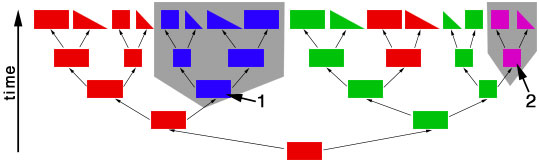 Arrow "1" points to the common ancestor of all blue objects (blueozoa) and arrow "2" points to the common ancestor of all pink objects (pinkozoa). The groups blueozoa and pinkozoa contain all descendants of the common ancestor. A group like this is called monophyletic, because they descend from a single ancestor, and it is called holophyletic, because it contains all descendants of this ancestor. These two groups are "groups of relatives" and thus "natural groups". But what about the other groups from our previous systems? Let us first consider the greenozoa: 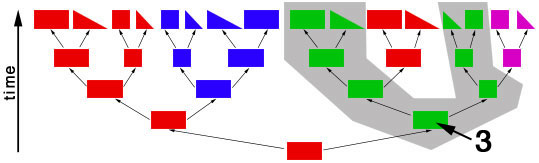 All greenozoa also derive from a single common ancestor (arrow "3"). The greenozoa are thus monophyletic. However, the greenozoa do not contain all descendants of this ancestor, because some of the red and all pink objects also derive from this ancestor. The greenozoa are thus not holophyletic. Groups that are monophyletic, but not holophyletic, are called paraphyletic. Let us now consider the redozoa: 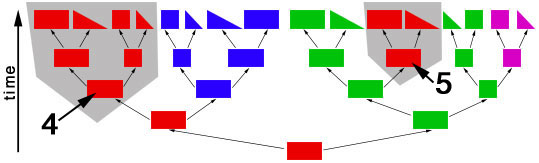 Originally we grouped these objects together, because they all share the character "red color". Now we see that not all red objects are directly related, because they derive from different ancestors (arrow "4" and arrow "5"). Thus, the redozoa is a group that contains unrelated objects characterized by a character that is identical in all objects, but that has evolved more than once. A group like this is called polyphyletic. All groups that we have formed by using the shape of the objects also are polyphyletic. The example below shows the distribution of the rectangulozoa in the tree: 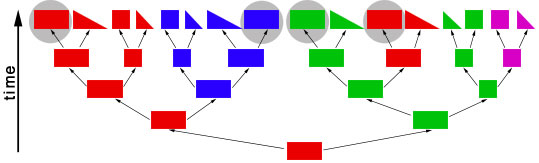 In summary, there are three different kinds of groups that we can form in the evolutionary tree: 1) monophyletic holophyletic groups (mostly simply called monophyletic groups) 2) monophyletic paraphyletic groups (mostly simply called paraphyletic groups) 3) polyphyletic groups All objects in the evolutionary tree are linked by descent. Only monophyletic holophyletic groups reflect this fact. Thus, for a natural system only monophyletic holophyletic groups are useful. We have seen that we obtained the desired monophyletic holophyletic groups only in some cases, depending on the character that we used. Thus, there are "good" and "bad" characters. A "good" character is e.g. "blue color", because it evolved only once in the ancestor of the blue group and is retained in all descendants. The blue color is thus an evolutionary innovation that occured in the ancestral blue object and still characterizes all extant objects of this ancestor. Such characters are called apomorphic characters or apomorphies. The apomorphy "blue color" characterizes the entire group (blueozoa). The scientific term for this is: the blue color is the autapomorphy for the blueozoa. But the blue color also characterizes each and every member of the blueozoa. The scientific term for this is: the blue color is the synapomorphy for the blue rectangle, blue square, blue triangle and the blue wedge. A character somewhat between "good" and "bad" is e.g. "rectangle shape". There are several objects with this character and these objects are not all directly related. Thus, this character leads to the polyphyletic group "rectangulozoa". Nevertheless the shape of these objects ultimately derives from a common ancestor that is also the ancestor of other objects: the rectangular shape is the ancestral state. Such a character is called plesiomorphic character or plesiomorphy. Plesiomorphies are normally useless for making groups. However, if a monophyletic group is already known based on other apomorphic characters, then plesiomorphies can nevertheless be useful to determine the direction of evolution of characters. Some characters evolve too quickly and can change back and forth frequently. In the example, shape is a character of this kind. These characters do not carry a phylogenetic signal and are useless for making groups, because these groups will be polyphyletic. Another character kind are characters that evolved in parallel in different branches of the tree. In the example, the red color is a character of this kind. The evolution of similar characters in different branches is called convergent evolution. The resulting similarities are called homoplasies. These characters also are useless for making groups, because these groups will also be polyphyletic. In practice, of course, apomorphies, plesiomorphies, and homoplasies can be difficult to identify. These problems are the cause for the many different hypotheses about metazoan phylogeny. A real example for a paraphyletic group is the Reptilia (reptiles). This group is based on characters like skin scales or the morphology of the heart, that once were believed to be apomorphic, but now are regarded as plesiomorphic characters. The ancestor of the Reptilia is also the ancestor of the birds (Aves). Thus the group Reptilia is monophyletic, but not holophyletic. A real example for a polyphyletic group is the Vermes (worms). This group is based mainly on the long thin shape of the body. This shape has evolved several times independently and is thus a homoplasy. |
| This page has been
updated on August 20, 2008 This site is online since May 31, 2005 Copyright © by Nikola-Michael Prpic. All rights reserved. |
Related information: |