

Ssp. lineatum:
(1) Cimex lineatus Linnaeus, 1758
Scutellara lineata (Linnaeus, 1758)
Scutellera lineata (Linnaeus, 1758)
Thyreocoris lineata (Linnaeus, 1758)
Tetyra lineata (Linnaeus, 1758)
Graphosoma lineata (Linnaeus, 1758) (Lapsus)
Graphosoma lineatum (Linnaeus, 1758)
Graphosoma lineatum lineatum (Linnaeus, 1758)
(2) Cimex nigrolineatus Fabricius, 1781
Tetyra nigrolineata (Fabricius, 1781)
Scutellera nigrolineata (Fabricius, 1781)
Trigonosoma nigrolineata (Fabricius, 1781)
Graphosoma nigrolineata (Fabricius, 1781)
(3) Graphosoma lineatum siciliensis Lupoli, 2017 (p. 25; TL: "Italie (Sicile), Castelbuono")
Ssp. italicum:
(1) Cimex italicus Müller, 1766 (p. 190; TL: "colliculum Cappucinorum, & secus fl. Duriam" (Norditalien))
Graphosoma lineatum italicum (Müller, 1766)
Graphosoma italicum (Müller, 1766)
Graphosoma italicum italicum (Müller, 1766)
(2) Tetyra schangini Gebler, 1817
(3) Scutellera rubrolineata Westwood, 1837
Graphosoma rubrolineata (Westwood, 1837) (Lapsus)
Graphosoma rubrolineatum (Westwood, 1837)
(4) Graphosoma flavipes Amyot, 1845 (nomen nudum)
(5) Graphosoma lineatum pallida Belke, 1859
(6) Scutellera crassa Motschulsky, 1861
Graphosoma crassa Motschulsky, 1861
(7) Scutellara lineata flavipes Mulsant et Rey, 1865
(8) Graphosoma lineatum reductum Horvath, 1901
(9) Graphosoma italicum ochraceum Royer, 1922/1923?
(10) Graphosoma italicum obscurum Stepanek, 1926
Graphosoma lineatum italicum obscura Stepanek, 1926
(11) Graphosoma italicum soosi Halaszfy, 1955
(12) Graphosoma italicum nigroscutellatum Halaszfy, 1955
(13) Graphosoma italicum flavolineata Halaszfy, 1955
Graphosoma italicum flavolineatum Halaszfy, 1955
(14) Graphosoma italicum sardiniensis Lupoli, 2017 (p. 25; TL: "Italie (Sardaigne), Santa Teresa")
(15) Graphosoma italicum nigriscutellatum auct. (Falschschreibung)
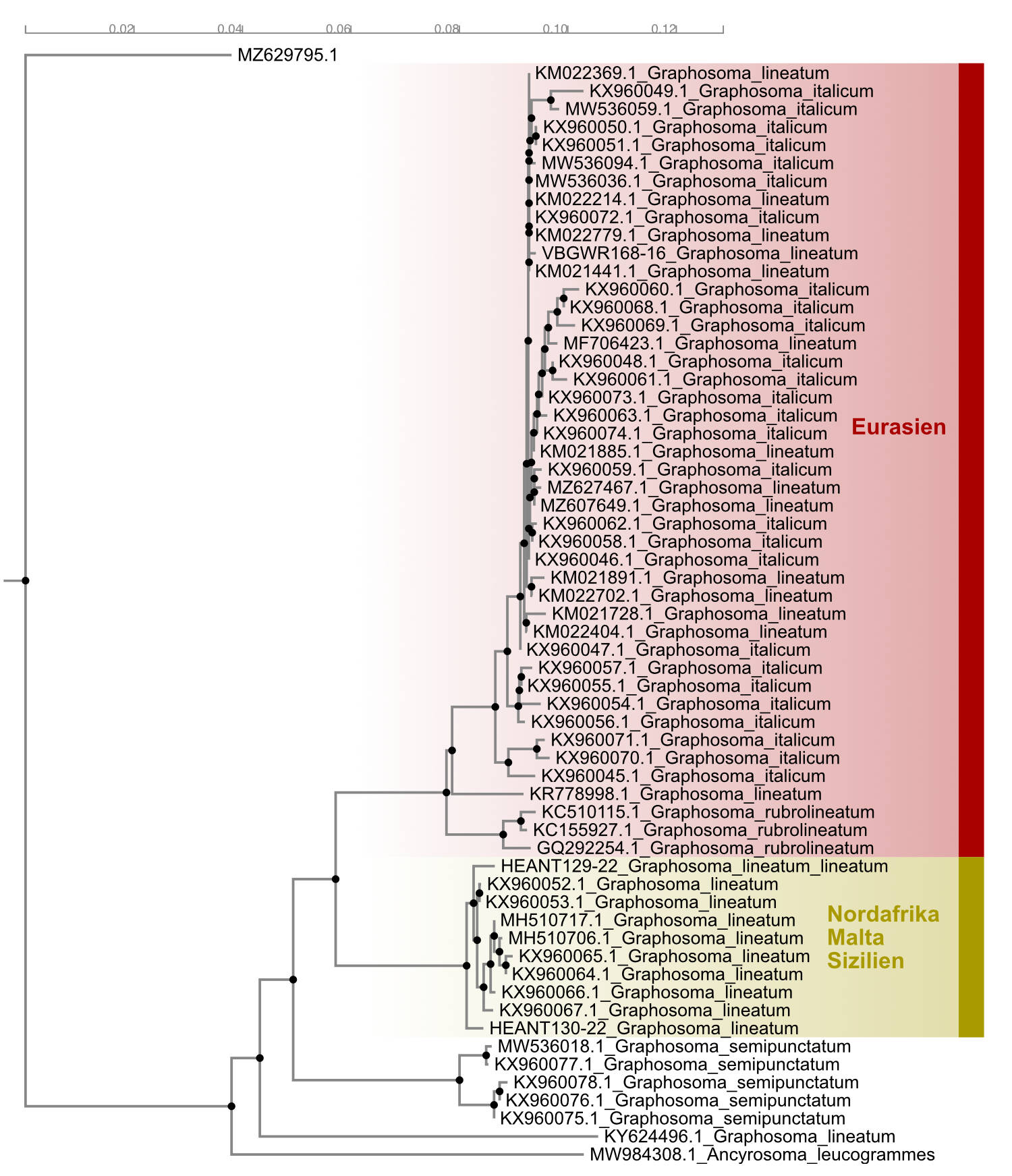
Es werden zwei Unterarten unterschieden: Graphosoma lineatum
lineatum in Nordafrika und den südöstlichen Mittelmeerinseln
Malta und Sizilien, und Graphosoma lineatum italicum im
restlichen Verbreitungsgebiet in ganz Europa und Asien.
Zur Systematik: die Populationen Nordafrikas werden auf
Grund von kleineren Färbungsdetails (vor allem der Beine)
traditionell von den Populationen des übrigen
Verbreitungsgebietes abgetrennt (Horvath 1903; Wagner 1956),
teils als Subspecies, teils als Species. Lupoli (2017) hat
beide Gruppen erneut in den Artrang erhoben. Diese
Einschätzung erfolgte auf der Basis der phylogenetischen
Analyse von Sequenzfragmenten des mitochondrialen Gens Mt-CO1
(dieses Gen codiert die Untereinheit I der
Cytochrom-c-Oxidase). Ich habe diese Analyse mit weiteren
inzwischen verfügbaren Sequenzfragmenten repliziert (siehe
Abb. 1). Die Ergebnisse sind denen von Lupoli (2017) ähnlich:
die Populationen Nordafrikas und der südlichen
Mittelmeerinseln Sizilien und Malta sind von den übrigen
Populationen deutlich getrennt. Die Interpretation dieser zwei
Gruppen als getrennte Arten ist jedoch nicht zwingend. Die
offenbar überlappungsfreie Allopatrie der beiden Gruppen
spricht zunächst nur für einen Unterartstatus. Es liegen keine
Untersuchungen zu Kontaktzonen oder (stabilen?) Hybridzonen
vor, die einen Artstatus wahrscheinlich machen könnten. Aus
meiner Sicht kann deshalb der Status der beiden Gruppen
derzeit nicht abschließend beurteilt werden und ich folge der
Argumentation von Pericart (2010), der die beiden Gruppen im
Rang von Subspecies führt. Die weiteren in Lupoli (2017)
beschriebenen Taxa sind unbegründet und gehören jeweils einer
der beiden Unterarten an. Auch das nominale Taxon Scutellera
rubrolineata ist Teil der eurasischen Unterart.
Zur Nomenklatur: die beiden Gruppen werden
üblicherweise als nominale Taxa Cimex italicus und Cimex
lineatus bezeichnet, da ersteres Taxon aus Norditalien und
letzteres Taxon aus Nordafrika beschrieben wurde. Damit
entsprechen die Typuslokalitäten zwar den auch genetisch
begründeten zwei Gruppen, die Taxa sind jedoch derzeit nicht
durch das entsprechende Typusmaterial auch nomenklatorisch
exakt bestimmt. Die Arbeit von Lupoli (2017) gibt zwar vor,
dieses Problem zu lösen, ist jedoch nomenklatorisch wertlos,
da für Cimex lineatus kein Typusmaterial untersucht wird und
für Cimex italicus unverständlicherweise ein "Lectotypus"
ausgewählt wird, obwohl das Typusmaterial verloren ist. Selbst
wenn man den "Lectotypus" stattdessen als Neotypus deutet,
entspricht die Wahl nicht den Erfordernissen, da der
"Lectotypus" nicht topotypisch ist: das Tier in Lupoli (2017)
stammt aus Saint-Martin-des-Londres in Frankreich, ein korrekt
gewählter, topotypischer Neotypus sollte jedoch an der
Typuslokalität ("colliculum Cappucinorum, & secus fl.
Duriam") in Norditalien gesammelt werden, also auf dem Turiner
Stadthügel Monte di Cappuccini und entlang des Flusses Dora
Riparia im nördlichen Stadtgebiet von Turin.
Deckert J, Wachmann E. 2020. Die Wanzen Deutschlands. Entdecken - Beobachten - Bestimmen. Quelle & Meyer Verlag, Wiebelsheim.
Horvath G. 1903. Conspectus specierum generis Graphosoma. Annales Musei Nationalis Hungarici 1, 345-354.
Lupoli R. 2017. Graphosoma lineatum (L., 1758) et G. italicum (O.F. Müller, 1766), deux espèces valides et distinctes, probablement issues de la transgression zancléenne méditerranéenne (Hemiptera Pentatomidae). L´Entomologiste 73, 19-33.
Madeira F, Madhusoodanan N, Lee J, Eusebi A, Niewielska A, Tivey ARN, Lopez R, Butcher S. 2024. The EMBL-EBI Job Dispatcher sequence analysis tools framework in 2024. Nucleic Acids Research 52(W1):W521-W525.
Müller OF. 1766. Manipulus insectorum Taurinensium. Melanges de Philosophie et de Mathematique de la Societe Royale de Turin 3, 185-198 (mit einem Vorwort von Carlo Allioni ("Carolus Allionius").
Pericart J. 2010. Hemipteres Pentatomoidea Euro-Mediterraneens. Volume 3. Systematique: Troisieme Partie Sous-Familles Podopinae et Asopinae. Faune de France 93, 1-291. Tafeln I – XXIV.
Ratnasingham S, Hebert PDN. 2007. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Molecular Ecology Notes 7, 355-364.
Sayers EW, Bolton EE, Brister JR, Canese K, Chan J, Comeau DC, Connor R, Funk K, Kelly C, Kim S, Madej T, Marchler-Bauer A, Lanczycki C, Lathrop S, Lu Z, Thibaud-Nissen F, Murphy T, Phan L, Skripchenko Y, Tse T, Wang J, Williams R, Trawick BW, Pruitt KD, Sherry ST. 2022. Database resources of the national center for biotechnology information. Nucleic Acids Research 50(D1):D20-D26.
Sievers F, Higgins DG. 2021. The Clustal Omega Multiple Alignment Package. Methods in Molecular Biology 2231, 3-16.
Wachmann E, Melber A, Deckert J. 2008. Wanzen. Band 4. Pentatomorpha II. In: Dahl F (Begründer). Die Tierwelt Deutschands, 81. Teil. Goecke & Evers, Keltern.
Wagner E. 1956. Systematik der Gattung Graphosoma Lap. (Hem., Het. Pentatomidae). Entomologische Berichten 16, 110-116.