Der Aufbau des unbefruchteten Vogeleies ist über alle
Vogelgruppen hinweg erstaunlich einheitlich (Abb. 1). Im
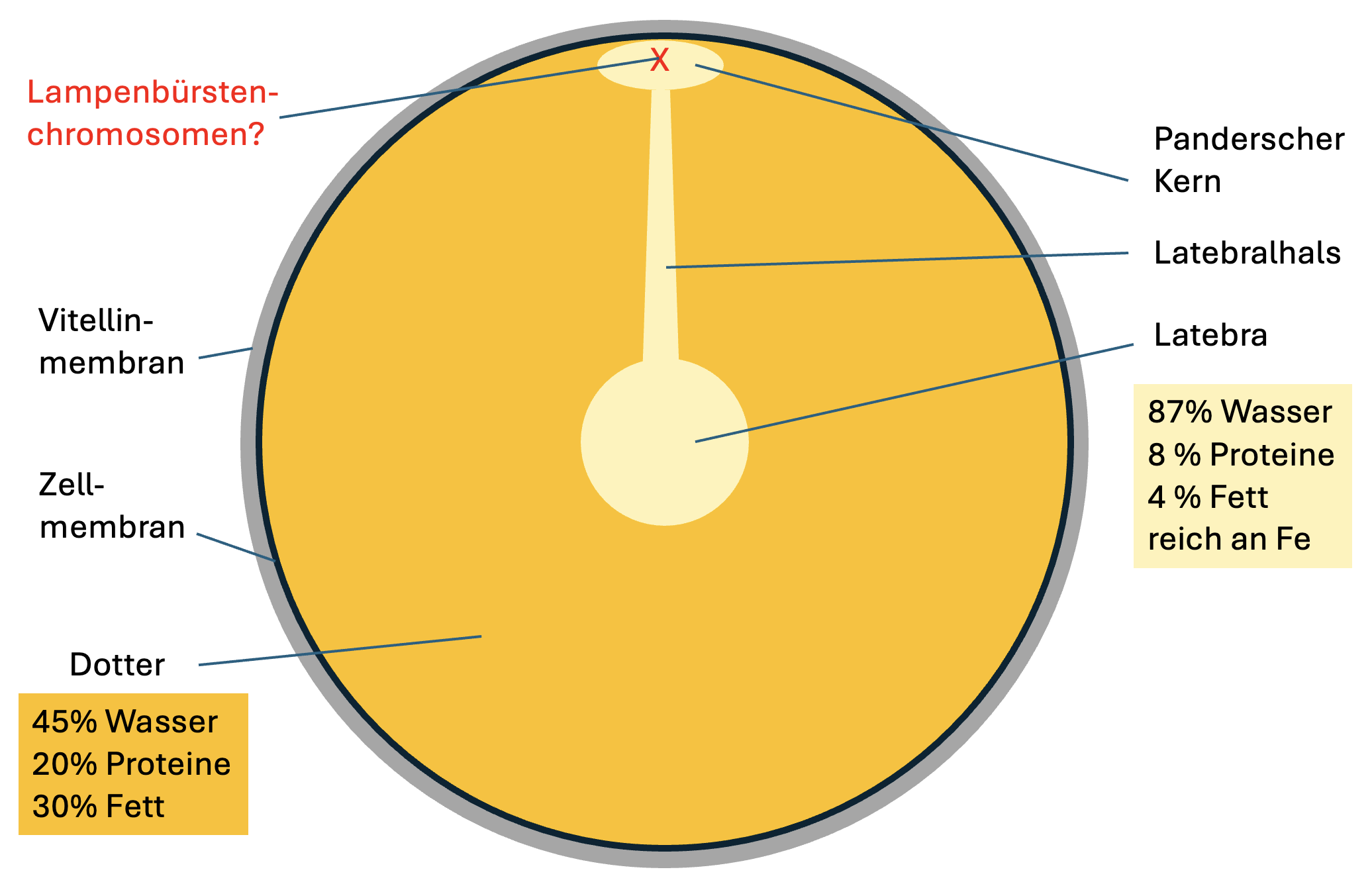
Zentrum des Eies liegt die Eizelle (Abb. 2). Wie jede Zelle so
besitzt auch die Eizelle eine Zellmembran. Auf der Oberfläche
der Zellmembran bildet die Eizelle eine dünne extrazelluläre
Matrix aus, die als Vitellinmembran ("Dottermembran")
bezeichnet wird. Die Eizelle nimmt während der Oogenese im
Ovar große Mengen an Dotter in ihr Cytoplasma auf. Dies
geschieht oft phasenweise, so dass der Dotter dann eine
konzentrische Schichtung aufweist. Die Schichten können sich
auch farblich unterscheiden, und werden dann als "weißer
Dotter" und "gelber Dotter" bezeichnet. Von den
Färbungsunterschieden abgesehen, die lediglich durch
Variationen der Pigmenteinlagerung zustande kommen,
unterscheiden sich der "weiße Dotter" und der "gelbe Dotter"
jedoch nicht in ihrer Zusammensetzung. Der eingelagerte Dotter
besteht zu etwa 45% aus Wasser, etwa 20% aus Proteinen und
etwa 30% aus Fetten. Nahezu das gesamte Cytoplasma der Eizelle
ist mit Dotter gefüllt. Lediglich in der Mitte befindet sich
ein kugelförmiger Bereich, die sogenannte Latebra, in dem das
Cytoplasma sehr wasserreich bleibt und kaum Proteine und Fette
enthält. Da dieser Cytoplasmabereich ebenfalls hell erscheint,
wird er manchmal ebenfalls irreführend als "weißer Dotter"
bezeichnet und darf dann nicht mit dem "weißen Dotter" im
eigentlichen Dotter verwechselt werden. Die Latebra setzt sich
in einen dünnen Strang fort, der als Latebrahals bezeichnet
wird, und der dicht unterhalb der Zellmembran in einen
ellipsenförmigen Bereich mündet, der als Panderscher Kern
(nucleus of Pander) bezeichnet wird (benannt nach Christian
Heinrich Pander (1794–1865)). Der Pandersche Kern ist
ebenso wie die Latebra und der Latebrahals ein wasserreicher
Bereich des Eizellen-Cytoplasmas, ist anders als Latebra und
Latebrahals jedoch im rohen Ei von außen als kleiner heller
Kreis an der Oberfläche des Dotters zu erkennen. Diese
durchscheinende Oberfläche des Panderschen Kerns wird oft als
"Keimfleck" oder fälschlich als "Keimscheibe" bezeichnet (die
echte Keimscheibe ist der frühe Embryo nach der Befruchtung).
In diesem Bereich entwickelt sich nach der Befruchtung dann
der Embryo; deshalb liegt es nahe, in dem wasserreichen
Cytoplasma des Panderschen Kerns in der unbefruchteten Eizelle
auch die in der Meiose I arretierten Lampenbürstenchromosomen
der Eizelle zu vermuten. Allerdings konnten Callebaut et al.
(2000) im Panderschen Kern weder mittels Feulgenscher Färbung
noch mittels Unna-Färbung Chromatin nachweisen.
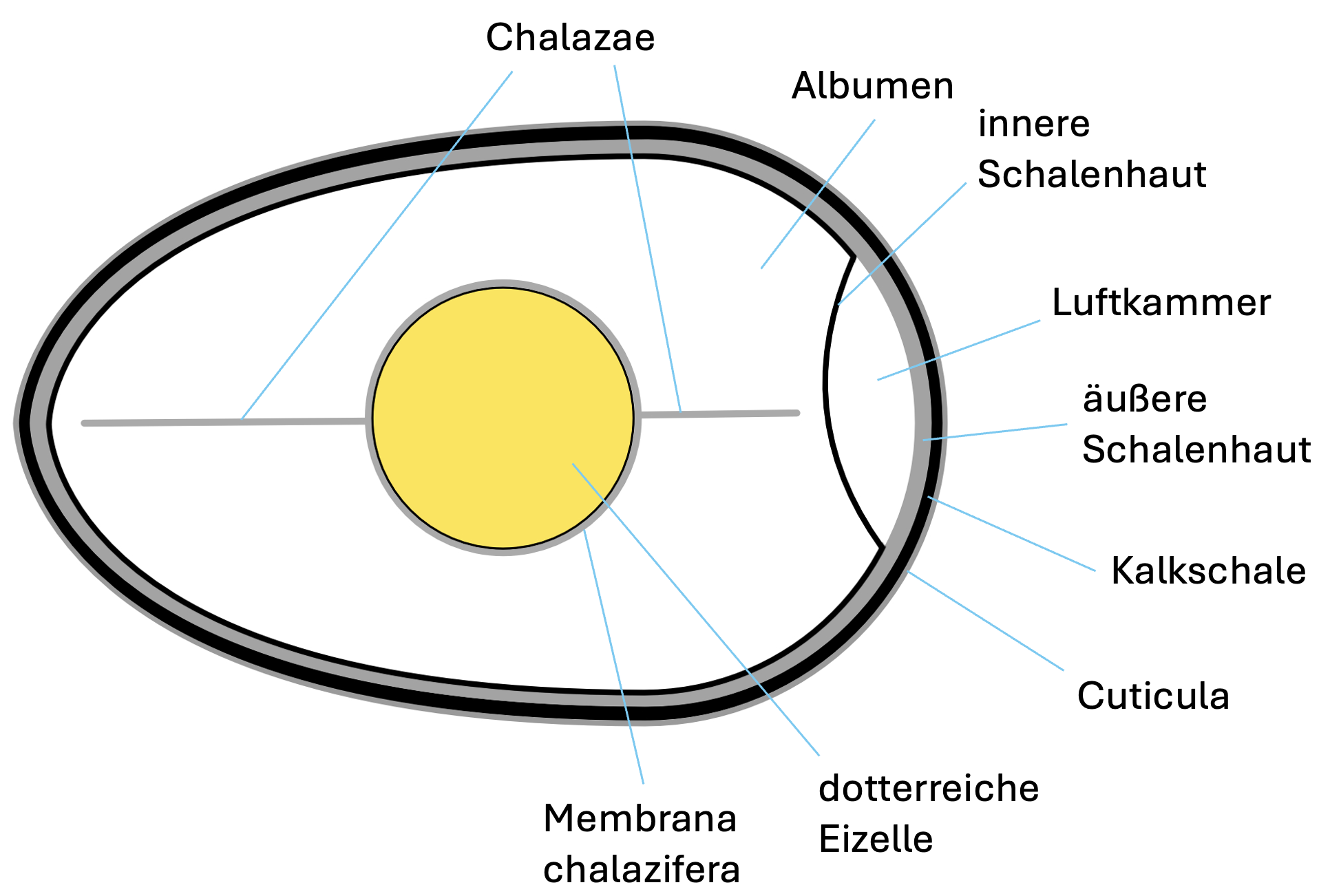
Im Eileiter wird die Eizelle mit den weiteren Bestandteilen
des Eies umhüllt und schließlich abgelegt. Im unbefruchteten
Ei sind dies:
(1) die Membrana chalazifera, eine Schicht aus sehr
zähflüssigem Ovalbumin, die in Richtung der beiden Eipole
jeweils in eine längliche Struktur ausläuft, die Chalaza
(Hagelschnur). Die beiden Chalazae verankern die Eizelle im
Albumen und helfen dabei die Position der Eizelle zu
stabilisieren, selbst bei starker Drehung des Eies. Die
Chalazae sind deshalb bei befruchteten und bebrüteten Eiern
meist vielfach verdreht, sorgen aber dafür, dass der Embryo
stets in der Eimitte und in Richtung zur Brutwärme der Eltern
zu liegen kommt.
(2) das Albumen ("Eiweiß", "Eiklar", "Weißei"), eine wässrige
Flüssigkeit, die vor allem die Proteine Ovalbumin,
Ovotransferrin (Conalbumin), Ovomucoid, Ovoglobuline, Lysozym
und Ovomucin enthält, daneben noch Spuren von Fetten und
weiteren Verbindungen und von Mineralstoffen. Das Albumen wird
in mehreren Etappen hinzugefügt und ist deshalb in drei Lagen
geschichtet, die sich auch bezüglich ihrer Viskosität stark
unterscheiden. Die dünne innere Lage, die auf die sehr zähe
Membrana chalazifera folgt, ist dünnflüssig. Darauf folgt eine
mächtige dickflüssige Lage und schließlich eine dünnere,
dünnflüssige Lage. Die Rolle des mengenmäßig in allen Lagen
weit überwiegenden Ovalbumins ist unbekannt, möglicherweise
wird es ähnlich wie der Dotter in der Eizelle für die
Versorgung des Embryos benötigt. Ovotransferrin, Ovomucoid,
die Ovoglobuline und Lysozym haben unter anderem starke
bakteriostatische Eigenschaften und schützen somit im
befruchteten Ei den Embryo vor Infektionen. Ovomucin ist ein
Glycoprotein, das wie andere Mucine durch seine Quellfähigkeit
die Viskosität der verschiedenen Albumenlagen bestimmt.
(3) die innere und die äußere Schalenhaut, die vor allem durch
ihren Gehalt an Kollagen, Keratin, Laminin und
Glukosaminoglykanen eine zähe und feste Schicht bilden. Beide
Schalenhäute sind fast überall mehr oder weniger stark
miteinander verschmolzen und bilden somit eine einheitliche
Schalenhaut. Nur am stumpfen Eiende (üblicherweise) weichen
die beiden Membranen auseinander und bilden zwischen sich die
Luftkammer.
(4) die mit Poren durchsetzte Kalkschale, aus mehreren,
verschieden aufgebauten Schichten, unter anderem einer
mächtigen Schicht aus Kalksäulen (Papillen).
(5) die Cuticula, eine feine Schicht mit wachsartiger
Konsistenz, die den Wasserverlust begrenzt, aber auch
zusätzlich vor dem Zutritt von Keimen in das Ei schützt.
Callebaut M, Van Nueten E, Harrisson F, Bortier H. 2000. Activation of avian embryo formation by unfertilized quail germ discs: comparison with early amphibian development. Reproduction Nutrition Development 40, 597-606.
Hutchison MJ, Lirette A, Etches RJ, Towner RA, Janzen EG. 1992. Research note: an assessment of egg yolk structure using magnetic resonance imaging. Poultry Science 71, 2117-2121.
King AS, McLelland J. 1978. Anatomie der Vögel. Verlag Eugen Ulmer, Stuttgart.