1. Erdmantelprozesse als wesentliche Ursache von
Erdoberflächenveränderungen
Die Entstehung der Erde aus Bestandteilen des sogenannten
"Sonnennebels" (eine Molekülwolke aus Gasen wie Wasserstoff
und Helium und kleineren Anteilen aus schwereren Molekülen)
hat vor etwa 4,6 Milliarden Jahren begonnen. Durch
Gravitationskräfte verdichtete sich der Sonnennebel zu
verschiedenen Himmelskörpern: gleichzeitig mit der frühen
Sonne (Protosonne) und der frühen Erde (Protoerde), bildeten
sich viele weitere Himmelskörper (Protoplaneten). Einige davon
sind noch heute erhalten (z. B. die Planeten, Monde,
Asteroiden und die transneptunischen Objekte), viele sind aber
in der Frühzeit der Bildung des Sonnensystems bei der
Kollision untereinander zerstört oder umgeformt worden. In
dieser frühen Phase der Entwicklung der Erde hat vermutlich
auch die Kollision eines solchen Protoplaneten (zuweilen
"Theia" genannt, nach der Mutter der griechischen Mondgöttin)
mit der Protoerde zur Bildung des Erdmondes geführt. Der Mond
hat durch seine Gravitationskräfte und seine Rotationszyklen
einen großen Einfluss auf die weitere Entwicklung der Erde und
die Evolution der Lebewesen genommen. Mit der Bildung der
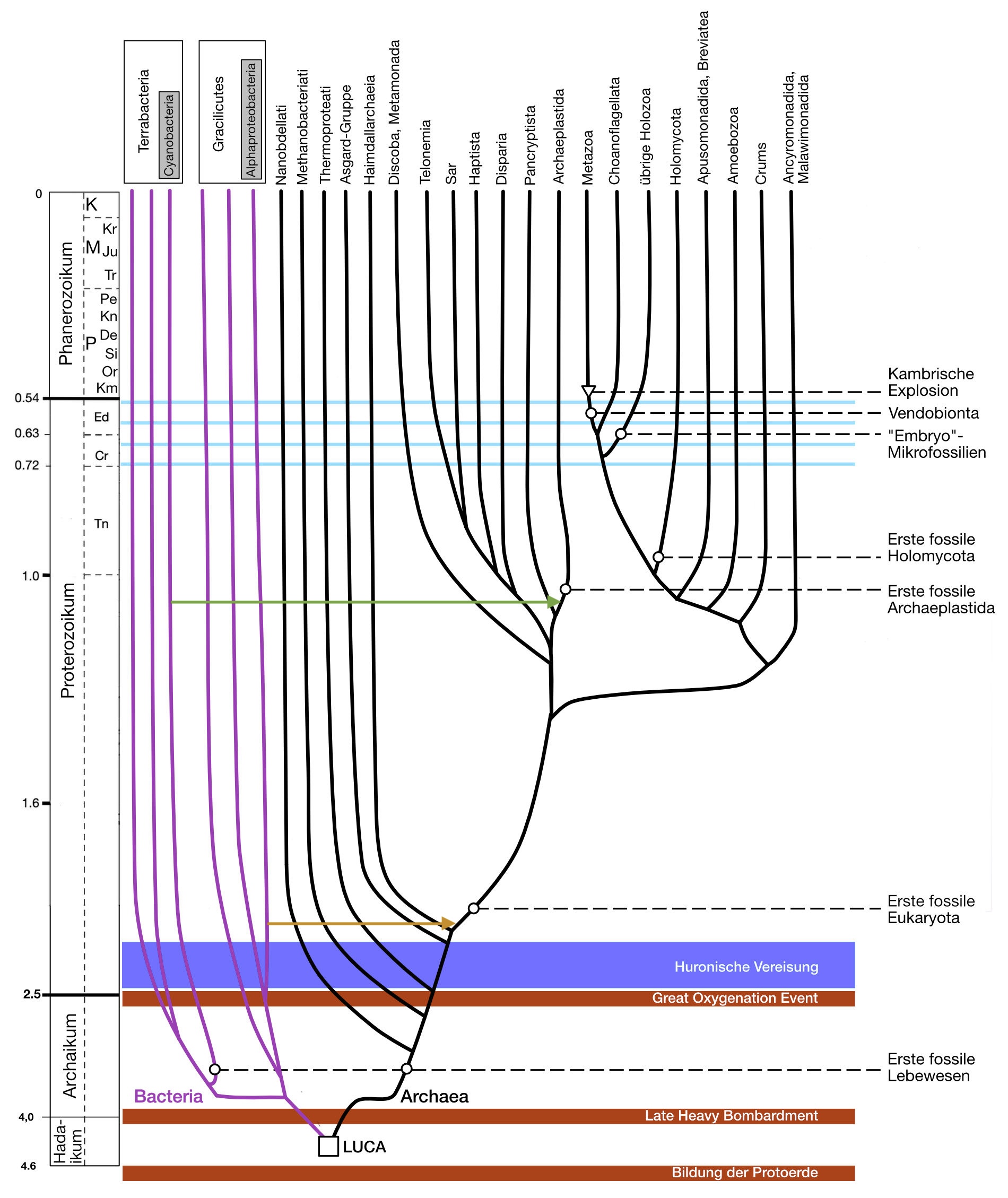
Protoerde beginnt auch die sog. geologische Zeitskala (Abb.
1), die in übersichtlicher Form die erdgeschichtlichen
Ereignisse seit der Erdentstehung bis heute darstellt.
Die Protoerde war zunächst sehr heiß, kühlte sich jedoch im weiteren Verlauf ab. Mit der Abkühlung entwickelte sich auch der Schichtaufbau der Erde, den wir heute feststellen können. Die äußerste Schicht, die Erdkruste, ist inzwischen so weit abgekühlt, dass sie erstarrt und fest ist. Unter der Erdkruste liegt der mächtige Erdmantel. Auch die sehr dünne äußerste Schicht des Erdmantels ist überwiegend erstarrt und bildet zusammen mit der Erdkruste die sogenannte Lithosphäre. Die weiter innen liegenden Anteile des Mantels sind jedoch plastisch verformbar und verhalten sich ähnlich wie eine Flüssigkeit. Die innerste Schicht des inneren Erdmantels (die sog. D''-Schicht) bildet den Übergang zum Erdkern. Der äußere Teil des Erdkerns besteht aus flüssigem Eisen, wohingegen der innere Erdkern wegen des hohen Drucks aus festem Eisen besteht, trotz der Temperaturen um 5700 °C, bei denen Eisen eigentlich flüssig ist. Die inneren Mantelbereiche werden vom heißen Erdkern erhitzt, kühlen jedoch mit zunehmender Nähe zur Erdoberfläche ab. Ganz ähnlich wie in einem Kochtopf auf der heißen Herdplatte kommt es dadurch zu ausgeprägten Konvektionsströmungen im plastischen Teil des Mantels. Das Mantelmaterial heizt sich in der Nähe des Kerns auf und steigt dadurch langsam in Richtung Lithosphäre auf. An der Lithosphäre wird die Strömung des aufsteigenden Mantelmaterials zu den Seiten abgelenkt und kühlt ab. Dadurch sinkt das Mantelmaterial nach einer Weile wieder in die Tiefe, wo es sich erneut erhitzt und danach erneut aufsteigt. Es bilden sich Kreisläufe von Konvektionsströmungen im Erdmantel. Durch diese Bewegung im Mantel kommt es zu Brüchen in der darüber liegenden, starren Lithosphäre: es entstehen einzelne, größere oder kleinere Lithosphärenplatten. Wo Plattengrenzen und aufsteigende Mantelkonvektion zusammentreffen, kann ein kleiner Teil des Mantelmaterials austreten, wird fest und fügt sich an die Lithosphärenplatten an. Dadurch wachsen die Platten an dieser Stelle und drücken den Rest der Platten weiter. Diese Schiebebewegung kann dazu führen, dass eine Lithosphärenplatte auf eine andere Lithosphärenplatte prallt. Am Ort des Zusammenpralls kommt es nun zur Subduktion: eine Platte wird unter die andere Platte geschoben (subduziert). Die subduzierte Platte durchstößt den oberen Mantel und dringt in den unteren Mantel ein. In einer Tiefe von ca. 1.000 Kilometern wird das Gestein eingeschmolzen und geht schlierenartig in der Konvektionsströmung im Mantel auf. Durch Prozesse wie den Austritt von Mantelmaterial und die Subduktion von Lithosphärenplatten stellen die Plattengrenzen die Kernorte von Erdbeben und Vulkanismus dar. Allerdings können beim Prozess der Subduktion Teile der geschmolzenen Platte sogar die Grenze zwischen Mantel und äußerem Kern erreichen und dort in der D''-Schicht neue Strömungen erzeugen, die als Mantel-Plumes oder Diapire bezeichnet werden. Diese gehören nicht zur bereits besprochenen zyklischen Mantelkonvektion, sondern steigen mehr oder weniger gerade nach oben auf und brechen dort durch die Lithosphäre. An diesen Stellen bilden sich sog. Hotspot-Vulkane. Hotspot-Vulkane sind also nicht an die Plattengrenzen gebunden, sondern können nahezu überall in der Lithosphäre auftreten; das bekannteste Beispiel sind die Vulkane des hawaiianischen Archipels, z. B. Mauna Loa und Kilauea inmitten in der pazifischen Platte.
Die komplexen Bewegungsprozesse in Kruste und Mantel tragen erheblich zur stetigen Veränderung der Erdoberfläche bei. Der Erdkern besteht zum größten Teil aus Eisen, wodurch ein ausgeprägtes Magnetfeld erzeugt wird. Auch dieses Magnetfeld wirkt auf die Prozesse der Umformung der Erdoberfläche ein. So werden beispielsweise neu gebildete Anteile der Lithosphäre während ihrer Bildung magnetisch beeinflusst und "speichern" dadurch ein regelmäßiges Muster aus sogenannten magnetischen Anomalien ab. An Bruchstrukturen wie z. B. dem mittelatlantischen Rücken tritt sehr langsam aber stetig flüssiges Gestein aus und verfestigt sich: die betroffenen Lithosphärenplatten wachsen und bewegen sich auseinander: der Meeresboden "wächst" (sog. Seafloor-Spreading). Das sich verfestigende Gestein erhält dabei eine Magnetisierung durch das Erdmagnetfeld. Da sich dieses im Mittel etwa alle 500.000 Jahre umpolt, wird im regelmäßigen Wechsel das neu austretende Material normal bzw. invers magnetisiert. Im Meeresboden entsteht somit ein verblüffend regelmäßiges "Streifenmuster" abwechselnder magnetischer Feldstärke, das z. B. Seevögeln und Walen zur Orientierung auf offener See dienen kann.
Die Verdichtung des Sonnennebels zur Protoerde und die anschließende Weiterentwicklung zur Erde mit dem soeben besprochenen Aufbau fanden im Hadaikum (auch: Hadeum) statt. Das Hadaikum bildet zusammen mit dem Archaikum und dem Proterozoikum den etwa 4,1 Milliarden Jahre dauernden Zeitraum vor dem Phanerozoikum (dem "Zeitalter des sichtbaren Lebens"; die Namensgebung fand statt, bevor man fossile Lebenspuren aus der Zeit vor dem Phanerozoikum kannte). Hadaikum, Archaikum und Proterozoikum werden geologisch in feinere Abschnitte unterteilt, die hier jedoch nicht weiter besprochen werden sollen. Dieser lange Zeitraum vor dem Kambrium wird vereinfachend als Präkambrium zusammengefasst.
2. Erste Lebewesen kurz nach der Entstehung der Protoerde
Da die Sequenzevolution der Nucleinsäuren im Erbgut der
Lebewesen inzwischen gut erforscht ist, ist es möglich durch
genetische Analysen der heute lebenden Organismen nicht nur
deren Verwandtschaft zu rekonstruieren, sondern den Stammbaum
auch zeitlich einzuordnen. Diese Berechnungen ergeben für den
gemeinsamen Vorfahr aller heute auf der Erde lebenden
Organismen (LUCA, last universal common ancestor) ein Alter
von etwa 4,3 Milliarden Jahren. Diese Ergebnisse sind
überraschend, denn sie bedeuten, dass es offenbar schon
300.000 Millionen Jahre nachdem sich die Protoerde aus dem
Sonnennebel gebildet hatte, bereits Leben auf der Erde gab.
Fossilien dieser ersten Lebewesen sind jedoch nicht bekannt.
Noch einmal etwa 300.000 Millionen Jahre später (vor ca. 4
Milliarden Jahren) kam es zum sogenannten Late Heavy
Bombardment, einer Phase vermehrter Meteoriteneinschläge im
gesamten Sonnensystem. Auf tektonisch aktiven Himmelskörpern
(wie z. B. der Erde), bei denen sich die Oberfläche
fortwährend verändert, sind die Spuren dieser Phase kaum mehr
sichtbar. Auf anderen, tektonisch nicht oder kaum aktiven
Himmelskörpern, wie z. B. dem Erdmond, dem Merkur oder einigen
Monden der Gasriesen, sind die Spuren des Late Heavy
Bombardments noch heute gut sichtbar: ihre Oberfläche ist mit
unzähligen Einschlagskratern übersät (Abb. 2). Wenn die
Berechnungen zum Alter von LUCA korrekt sind, dann müssen
Nachkommen von LUCA trotz der für das Leben denkbar
ungünstigen Bedingungen das Late Heavy Bombardment überlebt
haben. Tatsächlich geben seltene geologische Befunde, wie
beispielsweise besondere Eisenerze, die zwischen 4,3 und 4,0
Milliarden Jahre alt sind und wahrscheinlich von
eisenoxidierenden Bakterien stammen, Hinweise auf Leben vor
und während des Late Heavy Bombardments.

Indirekte Hinweise auf biologische Prozesse kurz nach dem Late Heavy Bombardment sind besondere Isotopenverhältnisse des Kohlenstoffs (Delta-C-13) in etwa 3,8 Milliarden Jahre alten Gesteinen. Die gefundenen Delta-C-13-Werte deuten auf photosynthetisch aktive Organismen hin, vermutlich Cyanobakterien. Aus etwa 3,8 bis 3,5 Milliarden Jahre alten Gesteinen kennt man außerdem Mikrofossilien, die von manchen Forschenden für fossile Bakterienzellen gehalten werden (Barghoorn & Schopf 1966; Schopf & Barghoorn 1967). Diese umstrittenen, etwa 1 bis 20 µm langen Strukturen wurden beispielsweise in Gesteinen aus Grönland (Barberton Greenstone-Gürtel), Südafrika (Fig Tree- und Onverwacht-Formation) und Australien (Warrawoona-Schichten) gefunden.
3. Fossile Stromatolithen: erste unzweifelhafte
Fossilien von Lebewesen
Die ersten unzweifelhaften Fossilien von Lebewesen finden
sich in Gesteinen mit einem Alter von etwa 3,5 Milliarden
Jahren. Es handelt sich dabei um fossile Stromatolithen
("geschichtete Steine"). Stromatolithen werden von
photosynthetisch aktiven Organismen im Wasser gebildet, indem
sie nach Überlagerung durch Sedimente versuchen, wieder an die
Oberfläche zu gelangen, um weiter dem Sonnenlicht ausgesetzt
zu sein (Reid et al. 2000). Durch diese über lange Zeiträume
wiederholten Wechsel von Sedimentüberlagerung und "Flucht" an
die Oberfläche, bilden sich regelmäßig und fein geschichtete
Gesteine, die Stromatolithen. Prinzipiell können diverse
photosynthetisch aktive Organismen Stromatolithen erzeugen, in
den allermeisten Fällen sind jedoch Cyanobakterien die
Stromatolithenbildner. Die Zellfäden der Cyanobakterien
wachsen an der Oberfläche der Stromatolithen, bis sie von
Sediment (z. B. feinem Sand, durch Wellen oder Strömungen
aufgewirbelt) bedeckt werden. Um weiter Photosynthese
betreiben zu können, wachsen die Zellfäden durch die neue
Sedimentschicht und bilden dort eine neue Lage. Die Zellen
unter der neuesten Sedimentschicht sterben ab. Auf diese Weise
lagern sich über lange Zeiträume immer neue Sedimentschichten
ab: der Stromatolith wächst in die Höhe. Fossile
Stromatolithen, die den heutigen Cyanobakterien-Stromatolithen
sehr stark ähneln, kennt man aus vielen erdgeschichtlichen
Perioden. Die ältesten bekannten fossilen Stromatolithen
stammen jedoch aus Schichten mit einem Alter von ca. 3,5
Milliarden Jahren und sind damit ein direkter Beleg für die
Existenz von Cyanobakterien im frühen Präkambrium (Allwood et
al. 2006). Ähnlich alt, nämlich 3,4 Milliarden Jahre, sind
fossile zelluläre und filamentöse Strukturen, die in der Nähe
von Hydrothermalquellen im Meeresboden nachgewiesen wurden
(Cavalazzi et al. 2021). Auf Grund ihrer chemischen
Zusammensetzung und ihres Vorkommens in einem methanreichen
Habitat werden diese Fossilien als älteste Nachweise von
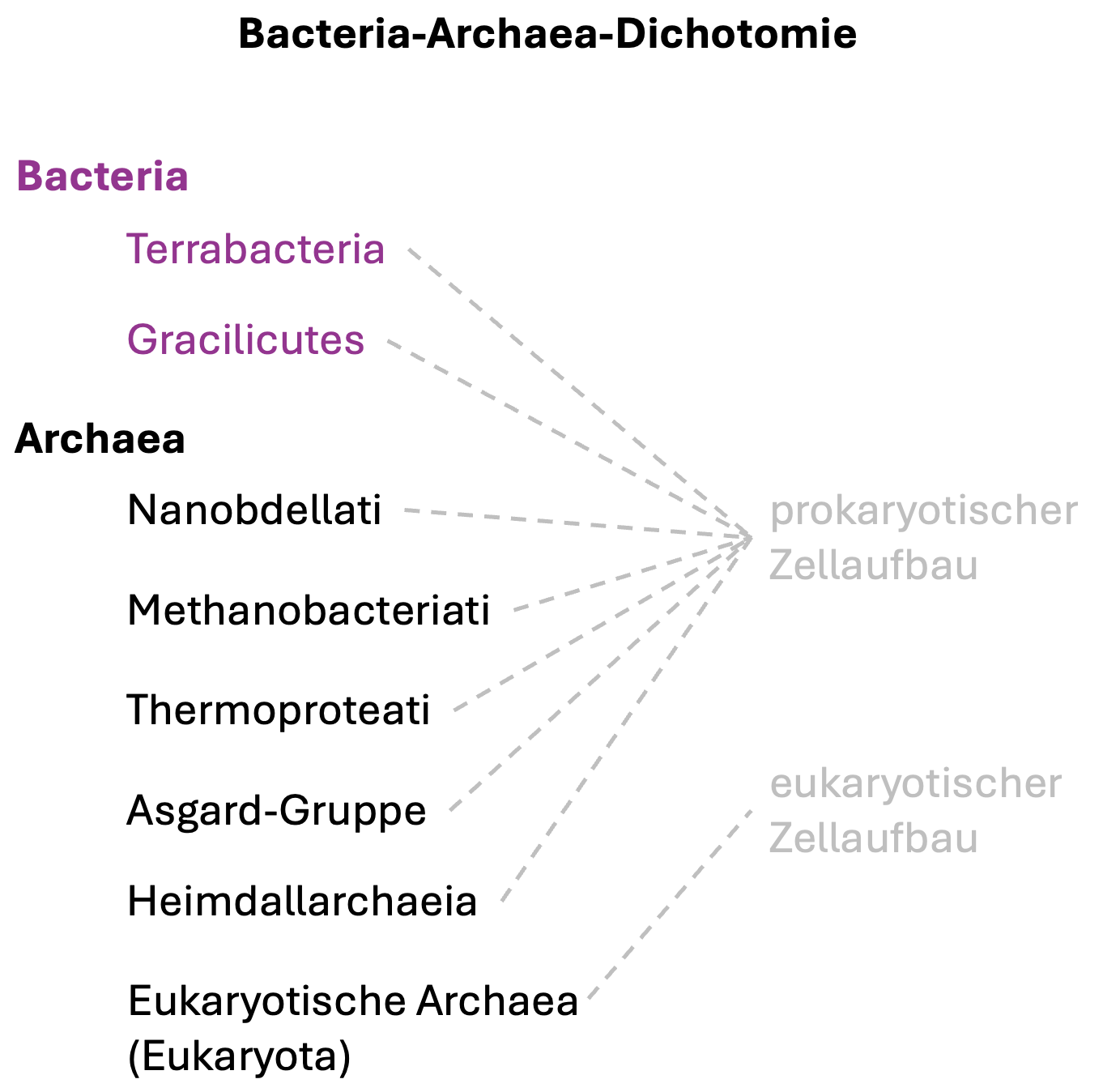
Archaea interpretiert. Somit hatte nach dem Late Heavy
Bombardment bereits die Aufspaltung in zwei getrennte
Abstammungslinien Bacteria und Archaea stattgefunden
(sogenannte Bacteria-Archaea-Dichotomie) (Abb. 3).
4. Die ersten eukaryotischen Lebewesen
Vor etwa 2,5 Milliarden Jahren kam es dann in direkter Folge
zu zwei tiefgreifenden Veränderungen, die sich geologisch
nachweisen lassen: dem Great Oxygenation Event (dem deutlichen
Anstieg des Sauerstoffgehalts in der Erdatmosphäre) und der
Huronischen Vereisung (der ersten globalen Vereisung).
Ursache, Dauer und Verlauf der beiden Prozesse sind
umstritten, es wird jedoch allgemein angenommen, dass sie
mindestens 300.000 Millionen Jahre andauerten. Für den Great
Oxygenation Event wird angenommen, dass Cyanobakterien, die zu
diesem Zeitpunkt bereits seit etwa 1 Milliarde Jahren
existierten und Photosynthese betrieben (deren Abfallprodukt
Sauerstoff ist), für den starken Anstieg des Sauerstoffgehalts
der Atmosphäre verantwortlich sind. Der Sauerstoffanstieg war
nicht kontinuierlich. Zeitweise war der Anstieg stetig und
gering. Zu anderen Zeiten gab es offenbar rapide Sprünge nach
oben, aber auch plötzliche Verringerungen der
Sauerstoffkonzentration. Auch die Huronische Vereisung
beinhaltet neben Phasen der vollständigen globalen Vereisung
("Snowball Earth") auch Phasen in denen zumindest Teile der
Erde nicht komplett vereist waren.
Da die Erdatmosphäre vor dem Great Oxygenation Event nahezu keinen Sauerstoff enthielt, hatten alle damaligen Lebewesen einen vollständig anaeroben Stoffwechsel. Sauerstoff behindert solche anaeroben Prozesse, wie z. B. die Reduktion von Eisenverbindungen, Schwefelwasserstoff und Methan. Das von den Cyanobakterien produzierte Photosynthese-Abfallprodukt Sauerstoff wirkte deshalb nicht nur für zeitgleich existierende nicht-photosynthetische Lebewesen giftig, sondern auch für die Cyanobakterien selbst. Es ist umstritten, wie die damaligen Lebewesen diese äußerst lebensfeindliche Phase der Kälte und Atmosphärenänderung überlebten. Es fällt aber auf, dass kurz nach dem Ende des Great Oxygenation Events und der Huronischen Vereisung die ersten eukaryotischen Lebewesen nachweisbar sind. Es liegt deshalb nahe, das Entstehen von Eukaryoten mit den lebensfeindlichen Umständen in Verbindung zu bringen.
Die ersten Eukaryoten, also die Lebewesen mit echtem Zellkern, sind fossil vor etwa 2,1 Milliarden Jahren nachweisbar. Die Fäden des Fossils Grypania sind im Gestein mit bloßem Auge zu erkennen. Sie sind mit einem Alter von 2,1 Milliarden Jahren die bislang ältesten bekannten Eukaryota-Fossilien. Einzellige und fädige Strukturen aus der Gunflint-Chert (USA und Kanada, knapp 2 Milliarden Jahre alt) sind überaus häufig. Ebenfalls aus der Gunflint-Chart sind erste sog. Acritarcha bekannt (González-Flores et al. 2022). Diese meist bizarr sternförmigen Mikrofossilien finden sich auch häufig in jüngeren Gesteinen, ihre Zuordnung zu einer Organismengruppe ist jedoch sehr umstritten. Umstritten ist auch die Einordnung der etwa 2 Milliarden Jahre alten, sackförmigen und kohlenstoffreichen Ablagerungen, die erstmals 1890 aus Finnland als Corycium enigmaticum beschrieben wurden. Mikroskopische Untersuchungen dieser Ablagerungen haben gezeigt, dass sie runde Strukturen beinhalten, die stark an eukaryotische Zellen mit einem Zellkern im Inneren erinnern (Matisto 1969).
Auch die molekulargenetischen Rekonstruktionen des Alters aller heute lebenden Eukaryoten ergeben ein ähnliches Alter. Somit fällt die Entstehung der Eukaryota mit großer Wahrscheinlichkeit auf die Zeit direkt nach dem Great Oxygenation Event und der Huronischen Vereisung. War die Evolution der komplexen eukaryotischen Zelle also tatsächlich eine Reaktion des Lebens auf die lebensfeindlichen Bedingungen in der Zeit vor etwa 2,5 bis 2,1 Milliarden Jahren? Neben der Frage nach der Entstehung des Lebens, ist auch die Frage nach der Entstehung der eukaryotischen Zelle auch heute noch unbeantwortet. Gesichert ist, dass es zu einer innigen Symbiose zwischen zwei verschiedenen prokaryotisch aufgebauten Zellen gekommen ist, und zwar zwischen einem Archaeon als Wirtszelle und einem Bakterium als Endosymbiont. Im Laufe der Zeit haben die Endosymbionten große Teile ihres Erbguts an den Wirt abgegeben, sind damit nicht mehr eigenständig lebensfähig und existieren als sogenannte Mitochondrien dauerhaft im Wirt. Die Mitochondrien werden in der Zellbiologie meist als Organellen der eukaryontischen Zelle aufgefasst, in Wirklichkeit aber sind es hochgradig angepasste endosymbiontische Bakterien. Sie verraten ihre Herkunft auch noch sehr gut, z. B. durch ihren bakterientypischen Aufbau, ihr eigenes Erbgut und eigene Ribosomen, und die Vermehrung durch inäquale oder äquale Querteilung. Auch genetisch sind sie noch als Bakterien zu erkennen und sind mit den Rickettsien (parasitäre Arten der Alphaproteobakterien) nah verwandt.
Auf die Entstehung der ersten Eukaryota-Zellen folgt eine längere Phase, in der weder im Fossilbericht noch durch molekulargenetische Studien eine Aufspaltung der Eukaryota in neue Abstammungsgemeinschaften erkennbar ist. Offenbar fand über einen Zeitraum von über 500 Millionen Jahren keine nennenswerte Diversifizierung der neu entstandenen Gruppe der Eukaryota statt. Erst vor etwa 1,5 Milliarden Jahren trennten sich die Eukaryota in zwei separate Abstammungsgemeinschaften auf, in die Diphoda und die Opimoda. Aber auch diese erste Diversifizierung (die sogenannte Diphoda-Opimoda-Dichotomie) blieb lange folgenlos, erst kurz vor Beginn des Toniums setzte in beiden Großgruppen die Bildung von zahlreichen getrennten Abstammungslinien ein. Innerhalb der Diphoda sind das die Gruppen Discoba, Metamonada, Telonemia, Sar, Haptista, Disparia, Pancryptista und Archaeplastida, und innerhalb der Opimoda sind das die Gruppen Ancyromonadida, Malawimonadida, Crums, Amoebozoa, Apusomonadida, Breviatea, Holomycota und Holozoa.
5. Eine photosynthetische Endosymbiose führt zur
Evolution der Archaeplastida
Bei einer Endosymbiose verfolgen Symbiont und Wirt in der Regel verschiedene Ziele. Der Symbiont erzielt durch das vollständige Eindringen in den Wirt eine weitgehende Unabhängigkeit von äußeren Umwelteinflüssen. Der Wirt dagegen ist meist auf den "Zugewinn" einer Fähigkeit des Symbionten aus. Bei der oben besprochenen Endosymbiose zwischen einem Archaeon und einem Alphaproteobakterium, die im Zusammenhang mit der Entstehung der eukaryotischen Zelle und ihrer Mitochondrien steht, stand für den Wirt vermutlich die Fähigkeit des Symbionten in Vordergrund, Sauerstoff in einem für den Wirt neuen Stoffwechselweg nutzen zu können. Eine weitere Eigenschaft, die für Wirte einen großen Vorteil bietet, ist die Fähigkeit Lichtenergie für den Stoffwechsel zu nutzen (Photosynthese). Endosymbiosen zwischen nicht-photosynthetischen Wirten und photosynthetischen Symbionten sind in der Natur deshalb überaus häufig. Die älteste dieser photosynthetischen Endosymbiosen ist die Aufnahme eines photosynthetisch aktiven Cyanobakteriums (als Endosymbiont) durch eine eukaryotische Diphoda-Zelle (als Wirt) vor etwa 1 Milliarde Jahren. Der Wirt war damit die erste eukaryotische Zelle, die Photosynthese betreiben konnte, und aus den photosynthetisch aktiven Endosymbionten sind im Laufe der Evolution die Plastiden (u. a. Chloroplasten) geworden. Alle Nachkommen dieser allerersten "Pflanzenzelle" bilden also eine geschlossene, vollständige Abstammungslinie, die als Archaeplastida ("die mit den ursprünglichen Plastiden") bezeichnet wird. Definiert man "Pflanzen" deshalb im engeren Sinne als alle Lebewesen, die von der ersten photoautotrophen Eukaryotenzelle abstammen, dann sind die Pflanzen in diesem Sinne eine natürliche Abstammungsgemeinschaft (monophyletische Gruppe). Die Archaeplastida sind also die Pflanzen im engeren Sinne.
Im weiteren Verlauf der Evolution haben sich andere Gruppen der Diphoda einzellige Archaeplastida als Endosymbionten zu Nutze gemacht und konnten so ebenfalls zu einer (größtenteils) photosynthetischen Lebensweise übergehen. Solche Endosymbiosen, in denen der Symbiont (also in diesem Fall einzellige Vertreter der Archaeplastida) bereits aus einer primären Endosymbiose hervorgegangen ist, nennt man sekundäre Endosymbiosen. Definiert man nun "Pflanzen" im weitesten Sinne als Lebewesen, die Photosynthese betreiben können, dann sind nicht nur die Archaeplastida "Pflanzen", sondern auch die verschiedenen durch sekundäre Endosymbiose zur Photosynthese befähigten Arten der Diphoda und auch die Cyanobacteria. In diesem Sinne sind "Pflanzen" keine natürliche Abstammungsgemeinschaft, sondern eine künstlich definierte Gruppe.
6. Präkambrische "Embryos": umstrittene Mikrofossilien
Im Verlauf der Erdentwicklung traten häufig Wechsel von Kalt- und Warmzeiten auf. Jedoch gab es nur wenige Phasen, in denen in der Kaltzeit tatsächlich eine globale Vereisung stattgefunden hat. Die Huronische Vereisung vor etwa 2,5 bis 2,2 Milliarden Jahren war die früheste und bei weitem längste dieser globalen Vereisungsphasen ("Snowball Earth"). Gegen das Ende des Präkambriums hin traten vier späte (und wesentlich kürzere) Snowball-Earth-Ereignisse ein. Die erste dieser späten Vereisungen ereignete sich im Zeitraum vor etwa 715 - 660 Millionen Jahren. Schon 10 Millionen Jahre später kam es zur zweiten Vereisung, vor etwa 650 - 635 Millionen Jahren. Die dritte und die vierte späte Vereisung waren beide vergleichsweise sehr kurz. Sie ereigneten sich vor etwa 579 Millionen Jahren und vor etwa 547 Millionen Jahren. Aus den Zeiträumen zwischen den globalen Vereisungen kennt man eine Reihe von Fossilien, die zeigen, dass die Lebewesen trotz der häufigen Vereisungsphasen die wärmeren Phasen dazwischen nutzen konnten, um sich auszubreiten und zu evolvieren.
Aus dem Zeitraum vor der ersten späten Vereisung sind sporen- und hyphenähnliche Fossilien bekannt, die als erste fossile Vertreter der Holomycota interpretiert werden (Loron et al. 2019). Aus dem Zeitraum zwischen der zweiten und dritten kürzeren Vereisung sind winzige Mikrofossilien (nur ca. 400 µm groß) bekannt, deren Form an Embryonalstadien verschiedener Tiere (im Sinne von Metazoa) erinnert. Diese Mikrofossilien sind deshalb anfänglich als "Embryo-Mikrofossilien" verschiedener Tierstämme bezeichnet worden (Chen et al. 2006; Yin et al. 2016; Yin et al. 2019; Xiao et al. 1998). Allerdings würde man dann erwarten, dass auch die fertig entwickelten Tiere im Fossilbericht dieser Fossillagerstätten enthalten sein sollten, was aber nicht der Fall ist: die größten Fossilien dieser Fundstellen sind nur etwa 1 bis 2 mm lang und erinnern nicht an bekannte Metazoa, sondern an die Zystenstadien von basalen Holozoa-Vertretern (Cunningham et al. 2016; Huldtgren et al. 2011). Die "Embryo-Mikrofossilien" sind also sehr wahrscheinlich wenigzellige Entwicklungsstadien im Lebenszyklus basaler Holozoa.
7. Rätselhafte Lebewesen des späten Präkambriums und
die Kambrische Explosion
Aus dem Zeitraum vor etwa 579 - 547 Millionen Jahren, also zwischen der dritten und der vierten späten Vereisung liegen dann wieder zahlreiche Fossilfunde vor, deren Interpretation ebenfalls stark umstritten ist. Allerdings sind die meisten davon recht groß (von wenigen Zentimetern bis hin zu über einem Meter), so dass man davon ausgeht, dass es sich überwiegend um "ausgewachsene" Organismen und nicht um Embryos oder andere Entwicklungsstadien handelt. Die Formen dieser Organismen sind recht vielgestaltig, allerdings ähneln die Lebewesen kaum irgendwelchen heutigen Arten, so dass es in der Fachwelt stark umstritten ist, worum es sich handelt. Es ist deshalb vorgeschlagen worden, diese Fossilien allgemein als "Vendobionta" zusammenzufassen, also wertungsfrei als "Lebewesen des Vendiums" zu bezeichnen (Vendium = veralteter Name des letzten Abschnitts des Präkambriums, heute Ediacarium genannt). Welche Rolle die Vendobionta bei der Entstehung der Metazoa gespielt haben, wird in der Forschung kontrovers diskutiert. Frühe Studien gingen davon aus, dass die Vendobionta keine Metazoa oder ihnen nah verwandte Lebewesen waren; es wurde diskutiert, ob es sich bei den Vendobionta z. B. um Verwandte von Pilzen oder verschiedenen Einzellergruppen handeln könnte. Aktuelle Interpretationen ordnen die Vendobionta jedoch überwiegend den Holozoa zu (und bezeichnen die Vendobionta deshalb als Ediacara-Fauna, also als "Tiere des Ediacariums"). Entweder sind die Vendobionta dann eine ausgestorbene Seitenlinie der Metazoa oder sogar die direkten Vorläufer der Metazoa. Letzteres würde auch den plötzlichen, explosiven Eindruck der "Kambrischen Explosion" deutlich relativieren (siehe unten). Tatsächlich befinden sich unter den späten fossilen Formen des Ediacariums bereits Arten, die unzweifelhaft zu den Metazoa gehören (Li et al. 2026).
Nach der Blüte der "Lebewesen des Vendiums" fand die vierte
späte Vereisung statt. Mit deren Ende vor etwa 547 Millionen
Jahren endet auch das Präkambrium, und die Periode des
Kambriums beginnt. In den kambrischen Gesteinsschichten findet
man erneut eine große Vielfalt an Fossilien. Diese Fossilien
ähneln aber heutigen Tiergruppen so sehr, dass kein Zweifel
daran besteht, dass es sich um echte, vielzellige Tiere (=
Metazoa) handelt. Es ist sogar sehr überraschend, dass man
Vertreter fast aller heutiger Tierstämme gefunden hat, es
waren also nicht nur basale Metazoagruppen, sondern auch schon
fast alle Stämme der Bilateria vertreten. Dieses erste
Auftreten der Metazoa ist also vor allem durch zwei
Besonderheiten gekennzeichnet:
(1) das Auftreten wirkt "plötzlich", weil es zuvor nur wenige
Vorläufer gab.
(2) das Auftreten wirkt "explosiv", weil offenbar in kürzester
Zeit eine riesige Vielfalt an Tiergruppen entstanden ist.
Dies hat zum etwas plakativen Begriff der "Kambrischen
Explosion" geführt.
Barghoorn ES, Schopf JW. 1966. Microorganisms three billion years old from the Precambrian of South Africa. Science 152, 758-763.
Cavalazzi B, Lemelle L, Simionovici A, Cady SL, Russell MJ, Bailo E, Canteri R, Enrico E, Manceau A, Maris A, Salomé M, Thomassot E, Bouden N, Tucoulou R, Hofmann A. 2021. Cellular remains in a ~3.42-billion-year-old subseafloor hydrothermal environment. Science Advances 7, eabf3963.
Chen JY, Bottjer DJ, Davidson EH, Dornbos SQ, Gao X, Yang YH, Li CW, Li G, Wang XQ, Xian DC, Wu HJ, Hwu YK, Tafforeau P. 2006. Phosphatized polar lobe-forming embryos from the Precambrian of southwest China. Science 312, 1644-1646.
Cunningham JA, Vargas K, Marone F, Bengtson S, Donoghue PC. A multicellular organism with embedded cell clusters from the Ediacaran Weng'an biota (Doushantuo Formation, South China). 2016. Evolution & Development 18, 308-316.
González-Flores AL, Jin J, Osinski GR, Tsujita CJ. 2022. Acritarch-like microorganisms from the 1.9 Ga Gunflint Chert, Canada. Astrobiology 22, 568-578.
Huldtgren T, Cunningham JA, Yin C, Stampanoni M, Marone F, Donoghue PC, Bengtson S. 2011. Fossilized nuclei and germination structures identify Ediacaran "animal embryos" as encysting protists. Science 334, 1696-1699.
Li G, Wei F, Wen W, Wang X, Lei X, Anderson RP, Zhao Y, Dunn FS, Parry LA, Cong P. 2026. The dawn of the Phanerozoic: a transitional fauna from the late Ediacaran of Southwest China. Science 392, 63-68.
Loron CC, François C, Rainbird RH, Turner EC, Borensztajn S, Javaux EJ. 2019. Early fungi from the Proterozoic era in Arctic Canada. Nature 570, 232-235.
Matisto A. 1969. On the microfossils of Corycium enigmaticum. Bulletin of the Geological Society of Finland 41, 199-202.
Reid RP, Visscher PT, Decho AW, Stolz JF, Bebout BM, Dupraz C, Macintyre IG, Paerl HW, Pinckney JL, Prufert-Bebout L, Steppe TF, DesMarais DJ. 2000. The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 406, 989-992.
Schopf JW, Barghoorn ES. 1967. Algal-like fossils from the Early Precambrian of South Africa. Science 156, 508-512.
Xiao S, Zhang Y, Knoll AH. 1998. Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite. Nature, 39, 553-558.
Yin Z, Zhu M, Bottjer DJ, Zhao F, Tafforeau P. 2016. Meroblastic cleavage identifies some Ediacaran Doushantuo (China) embryo-like fossils as metazoans. Geology 44, 735-738.
Yin Z, Vargas K, Cunningham J, Bengtson S, Zhu M, Marone F, Donoghue P. 2019. The Early Ediacaran Caveasphaera Foreshadows the Evolutionary Origin of Animal-like Embryology. Current Biology 29, 4307-4314.