
Fig. 1: Fig. 1. Female imago of the Common Lacewing Wasp Helorus anomalipes. After Pschorn-Walcher (1971).
The Heloridae are small parasitic hymenopterans (see Fig. 1 for an example). The biology of the family is insufficiently known and the species boundaries are debated even for the species of the German fauna. In the older literature the different species are often lumped together into one single species or the different species are misidentified. Thus, further research on the biology of the Heloridae is still an interesting and rewarding field of study.
The Heloridae currently comprise only the single genus Helorus containing about 10 species worldwide. The majority of species occurs in the Palearctic region, but some species are also known to occur in south and central America, Africa, and the Indo-Australian region. The German representatives of the family are mainly black and only the legs can be partially yellow or red. A characteristic trait of the Heloridae are the combed claws on the tarsus, but this can only be seen at high magnification and this trait is not suitable for field identification. Like all representatives of the parasitic wasps of the superfamily Proctotrupoidea the Heloridae have reduced wing venation. However, compared to other proctotrupoids the Heloridae retain a relatively well-developed venation in their forewings, which is highly characteristic for the family (Fig. 2). The shape of the pterostigma, that is formed by the veins at the anterior margin, is important for species identification. The venation of the hindwings is almost completely reduced except for the vein along the anterior margin and a weakly sclerotized crossvein.
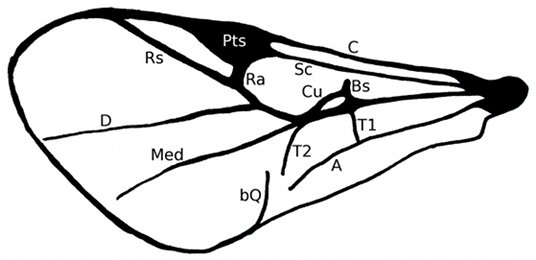
Fig. 2: Forewing venation of the Heloridae.
The wing root gives rise to several radial veins, namely the analis (A),
costa (C) that runs around the entire wing, subcosta (Sc) that terminates at
the pterostigma (Pts), and the medialis (Med). The medialis gives rise to the
short veins cubitalis (Cu) and basalis (Bs), and the transversal veins T1 and
T2 and the radialis (Rs). The radialis gives rise to the short radial branch (Ra)
and the discoidalis (D). The so-called curved crossvein (bQ) is a weakly sclerotized,
blind vein at the posterior margin. Apart from this vein classification scheme there
are several other systems used in the literature that are based on different ideas
about vein evolution and homology. These are not used here. After Pschorn-Walcher (1971).
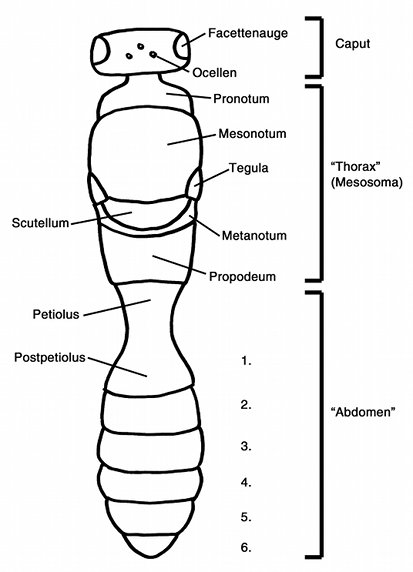
Fig. 3: Schematic overview of the body plan of
higher hymenopterans. The anterior trunk comprises the thorax proper and the
first abdominal segment (propodeum) and is therefore properly called mesosoma.
However, most authors nevertheless use the term "thorax" and consequently
the numbering of the abdominal segments begins not with the propodeum, but
with the next abdominal segment. Usually 6 abdominal segments are visible in
dorsal aspect, but differences can exist between males and females. Additional
modifications exist due to tergite fusions (e.g. in Heloridae).
The thorax proper of the Heloridae is dorsally covered by a small pronotum and a large mesonotum, which has a number of deep furrows (prescutal sutures or notaulices). The scutellum is a small and oval plate that has a species-specific sculpture. The head (caput) of the Heloridae is almost rectangular in shape and bears the mouthparts and the antennae. The antennae consist of 16 segments (including a small ring (anellus) at the base of the flagellum that is often not considered to be a "true" segment). The relative length of these antennal segments is sometimes used for species identification, but this is problematic, because in most species the intraspecific variation of these traits is not known.
As mentioned above, the biology of the species is insufficiently known. The females inject their eggs into the larvae of lacewings; obviously only a single egg per larva is deposited. The first larval instar of the wasp waits until the pupation and cocoon formation of the host, before killing the host. The following larval stages feed on the host tissue und the final larval instar drills a hole into the host cocoon. However, it does not leave the cocoon entirely but remains with its abdomen in the cocoon and also pupates in this position. The imago ecloses after 8 to 12 days and the full life cycle takes about 30 days (under laboratory conditions). The imagines are primarily found in April to October (with a peak in the summer), but the number of generations during this time is not known. The phenology differs between species and it is possible that species with the same host species alternate to avoid damage to the host population; however, host specificity of the Helorus species is not known so far. For some species only a single lacewing species is known, but for other species several species have been reported. For Helorus anomalipes a species of Hemerobius is also reported, but this is very likely based on a misidentification of the parasite species. All reliable host records are exclusively refering to lacewings. The food source of the imagines is not known, but there is a report of Helorus anomalipes female imagines visiting flowers of Peucedanum oreoselinum (Hedicke 1922), suggesting that they may feed on nectar or pollen. The ocurrence of the species of course depends on the distribution of the host species; it is unclear whether other ecological factors play a role as well.