Vielzelligkeit als Merkmal der Metazoa
Die monophyletische Gruppe der Metazoa umfasst diejenigen
Lebewesen, die wir ohne weiteres als "Tiere" ansprechen
würden. Ein wichtiges Merkmal der Metazoa ist ihre
Vielzelligkeit. Allerdings ist das kein spezifisches Merkmal
der Metazoa: die Vielzelligkeit ist während der Evolution sehr
oft entstanden. Fast alle größeren monophyletischen Gruppen
der Eukaryota haben unabhängig die Vielzelligkeit erreicht. In
der Gruppe der Excavata sehen wir bei den Fornicata zwar noch
keine echte Vielzelligkeit, aber eine besondere Form der
"Zweizelligkeit" (z. B. Gattung Giardia), die vermutlich
dadurch erreicht wird, dass eine normale Zellteilung begonnen
wird, aber nicht vollständig abgeschlossen wird. Bei den
Discoba gibt es aber schon mehrzellige Kolonien in der Gruppe
der Euglenozoa, und auch Vielzelligkeit bei Arten der
Heterolobosea, z. B. die "Schleimpilze" der Gattung Acrasis.
In der Gruppe der Diaphoretickes ist bei den Pancryptista
zumindest eine wenigzellige Art der Cryptophyta bekannt
(Bjornbergiella hawaiiensis) und koloniale Formen gibt es bei
den Centrohelida (z. B. Raphidiophrys viridis) und den
Alveolata (z. B. Gattung Zoothamnium), netzwerkbildende Formen
bei den Rhizaria (z. B. Gattung Chlorarachnion) und eine große
Vielfalt an kolonialen bis vielzelligen Formen bei den
Stramenopila (z. B. die Eipilze und die Braunalgen). Die
Stramenopila haben die Vielzelligkeit daher mit Sicherheit
sogar mehrfach evolviert. Das gleiche gilt vermutlich auch für
die Archaeplastida, wo wir unterschiedliche Vielzellertypen z.
B. bei den Grünalgen sehen, und natürlich richtige Vielzeller
sowohl bei den Rotalgen als auch bei den grünen Landpflanzen
finden. Auch innerhalb der Amorphea, mit ihrer immensen
Vielfalt an kolonialen und wenigzelligen Formen und den
verschiedensten echten Vielzellern, wurde die Vielzelligkeit
mit großer Wahrscheinlichkeit mehrfach evolviert. Neben den
vielzelligen Tieren (Metazoa), finden wir Vielzelligkeit auch
bei den Amoebozoa (vor allem bei den "Schleimpilzen" der
Gattung Dictyostelium), den Holomycota ("Pilze") und auch bei
ursprünglichen Gruppen der Holozoa selbst (z. B.
Choanoflagellata).
Es gibt also viele evolutionäre Wege, die zur Vielzelligkeit
führen und diese Wege wurden auch mehrfach unabhängig
voneinander beschritten. Das zeigt sich besonders gut daran,
dass nicht nur eukaryotische Organismen die Vielzelligkeit
erreicht haben, sondern dass es auch bei den Bacteria
vielzellige Gruppen gibt (z. B. Gattungen Nostoc, Fischerella,
Schizothrix). Am beeindruckendsten ist vielleicht die
mehrfache Evolution der Bildung vielzelliger Körper durch
Aggregation unabhängiger Einzelzellen. Die Ähnlichkeit dieses
Vorgangs bei so unterschiedlichen Gruppen wie Myxobakterien,
Heterolobosea und Amoebozoa ist ein ganz außergewöhnlicher
Fall von konvergenter Evolution.
Besonderes Epithelgewebe als apomorphes Merkmal der Metazoa
Die Vielzelligkeit bei den Metazoa ist jedoch die Grundlage
für eine Apomorphie der Metazoa, nämlich die Ausbildung eines
besonderen Typs von Epithelgewebe. Da Gewebe immer aus
mehreren Zellen bestehen, ist die Vielzelligkeit natürlich
eine Voraussetzung, um auch Epithelgewebe bilden zu können.
Jedoch sind nicht alle vielzelligen Zusammenschlüsse auch
gleich Epithelien. Wie schon die Vielzelligkeit, so ist auch
das Vorhandensein von Epithelien kein exklusives Merkmal der
Metazoa, denn z. B. die hoch organisierten Viridiplantae
(grüne Pflanzen) haben unabhängig von den Metazoa ebenfalls
Epithelien evolviert. Allerdings ist der morphologische Aufbau
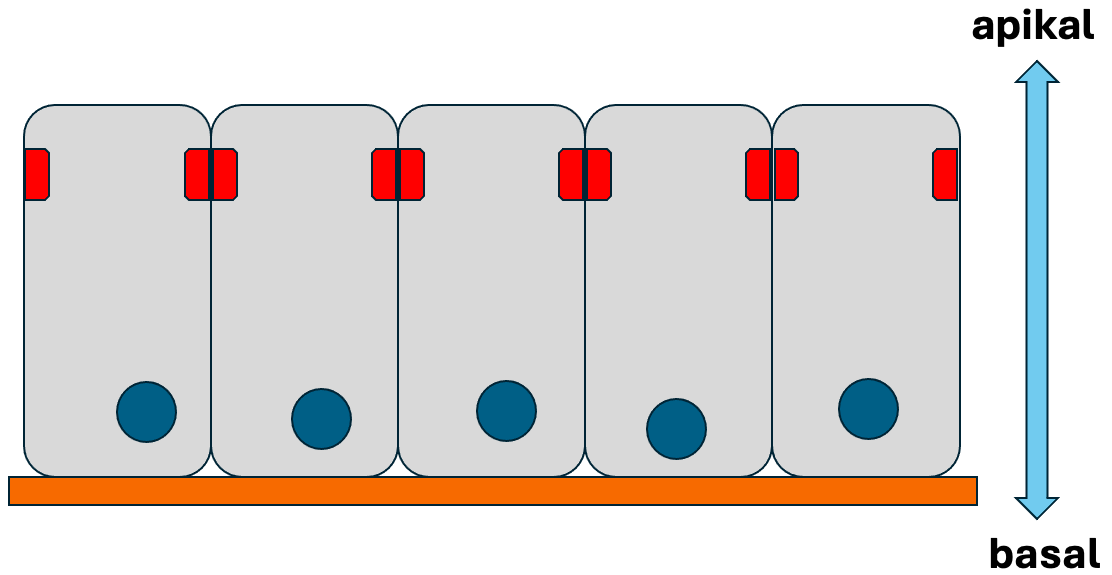
der Epithelien der Metazoa sehr charakteristisch und findet
sich in dieser Form nur bei den Metazoa. Vereinfacht
beschrieben, sind metazoische Epithelien geschlossene
Zellverbände, deren Zellen auf einer gemeinsamen basalen
Matrix aufsitzen und am gegenüberliegenden Zell-Ende durch
Verschlusskontakte fest miteinander verbunden sind. Die Zellen
in einem Epithel haben also stets eine deutliche Polarität mit
einem basalen Ende (in Richtung der basalen Matrix) und einem
apikalen Ende (in Richtung der Verschlusskontakte). In Abb. 1
ist ein stark vereinfachtes Schema eines metazoischen Epithels
dargestellt.
Das erste (primordiale) Epithel eines Individuums entsteht
bereits in der Frühphase der Embryonalentwicklung. Aus der
befruchteten Eizelle (Zygote) entstehen durch Zellteilungen
(Furchungsteilungen) zunächst mehrzellige Stadien
(2-Zell-Stadium, 4-Zell-Stadium usw.), die jedoch noch nicht
gewebeartig organisiert sind. Durch weitere Zellteilungen
bildet sich aber schon kurze Zeit später eine vielzellige,
innen üblicherweise hohle "Zellkugel", die Blastula. Die
Zellen, die die Blastula bilden, sind nun als Epithel
organisiert. Dieses primordiale Epithel wird als Blastoderm
bezeichnet (Abb. 2 links). Die Zellen des Blastoderms
differenzieren sich weiter: es entstehen die primären
Keimblätter. Der Name ist unglücklich gewählt, da eine
Verwechslung mit den Keimblättern bei den Pflanzen droht. Die
Keimblätter der Metazoa haben mit den pflanzlichen
Keimblättern jedoch überhaupt nichts zu tun. Der Name
"Keimblätter" soll bei den Metazoa stattdessen zwei Punkte zum
Ausdruck bringen:
(1) es handelt um Epithelien. Für den Fachbegriff "Epithelium"
war im 19. und frühen 20. Jahrhundert der deutschsprachige
Begriff "Blatt" üblich, was aber nichts mit den Blättern bei
Pflanzen zu tun hat.
(2) sie sind Quelle bzw. Ausgangspunkt für die Entwicklung von
Organen. Auch hier ist wieder der Begriff "Keim" eine alte
Formulierung für "Quelle, Ausgangspunkt".
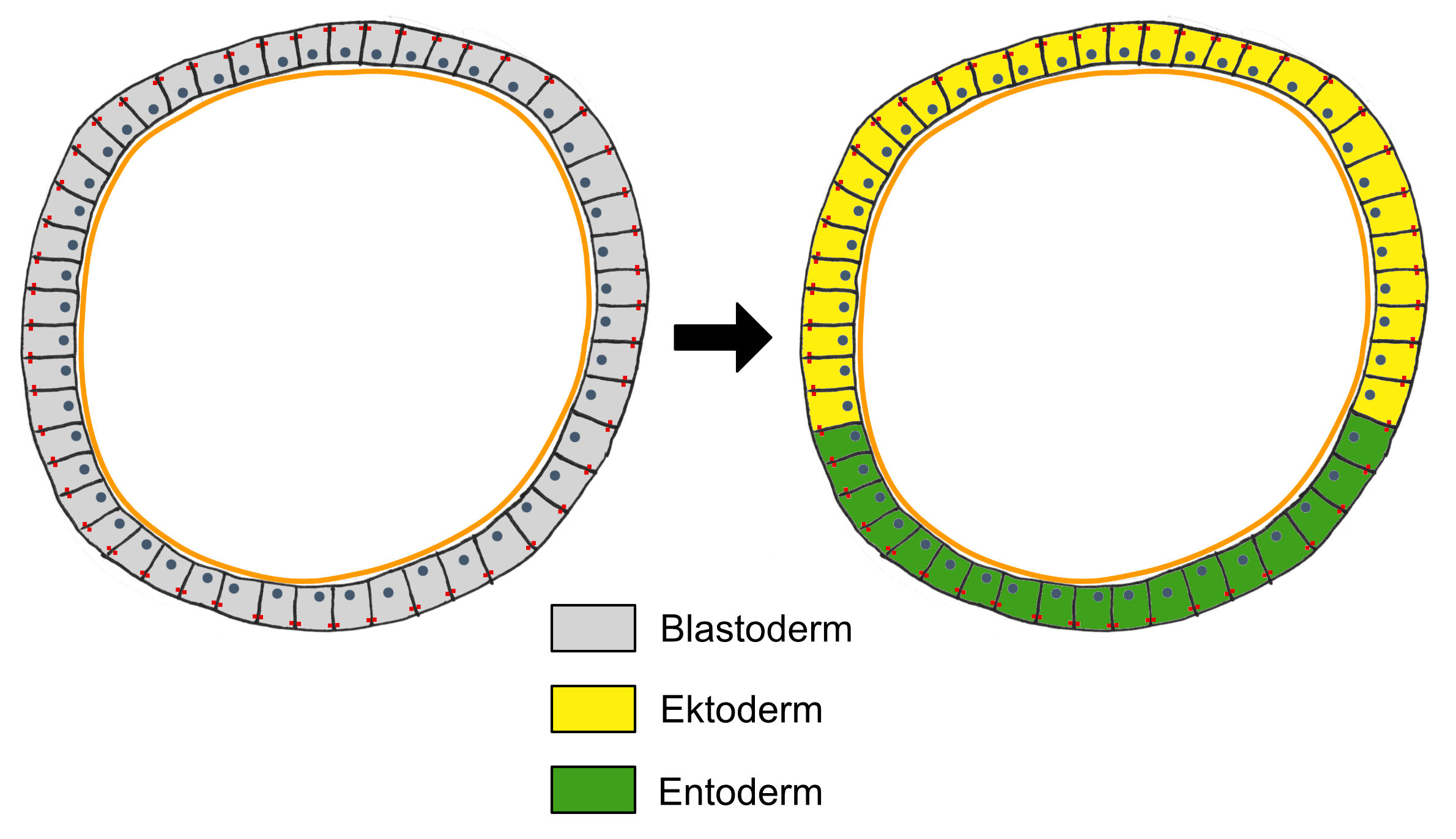
Die Differenzierung des Blastoderms in die primären
Keimblätter findet meist noch im Blastulastadium statt. Ein
Teil der Blastodermzellen erhält dabei ein ektodermales
Schicksal (gelb in Abb. 2, rechts), der andere Teil der
Blastodermzellen erhält stattdessen ein entodermales Schicksal
(grün in Abb. 2, rechts). Danach setzen umwälzende embryonale
Entwicklungsmechanismen ein, die dafür sorgen, dass die
determinierten Zellen an ihren vorgesehenen Bestimmungsort
gelangen und sich differenzieren können: die ektodermalen
Zellen bleiben zunächst an der Oberfläche, während die
entodermalen Zellen in das Innere der Blastula gelangen.
Dieser Prozess wird als Gastrulation bezeichnet, sein Ergebnis
ist die Gastrula, die nun aus den beiden primären Keimblättern
Ektoderm (außen) und Entoderm (innen) besteht. Meistens bildet
sich während der späten Phase der Gastrulation aus Teilen der
bereits vorhandenen primären Keimblätter ein weiteres,
sekundäres Keimblatt, das als Mesoderm bezeichnet wird.
Für die Bildung des Mesoderms gibt es diverse Mechanismen,
prinzpiell stammen aber die mesodermalen Zellen ursprünglich
entweder aus dem Ektoderm oder aus dem Entoderm: man spricht
differenzierend von Ektomesoderm bzw. Entomesoderm. Beides
schließt sich aber nicht gegenseitig aus, so dass in einer Art
sowohl Ekto- als auch Entomesoderm vorliegen kann. Die Zellen
des Mesoderms stammen also aus Epithelien und sind somit mit
der Fähigkeit Epithelien zu bilden vollständig ausgestattet.
Nicht immer wird diese Fähigkeit jedoch auch umgesetzt. Neben
epithelial organisiertem Mesoderm (z. B. als Cölothelien um
sekundäre Leibeshöhlen (sog. Cölome)) bilden mesodermalen
Zellen aber auch andere Gewebetypen, allen voran das
Bindegewebe und das Muskelgewebe.