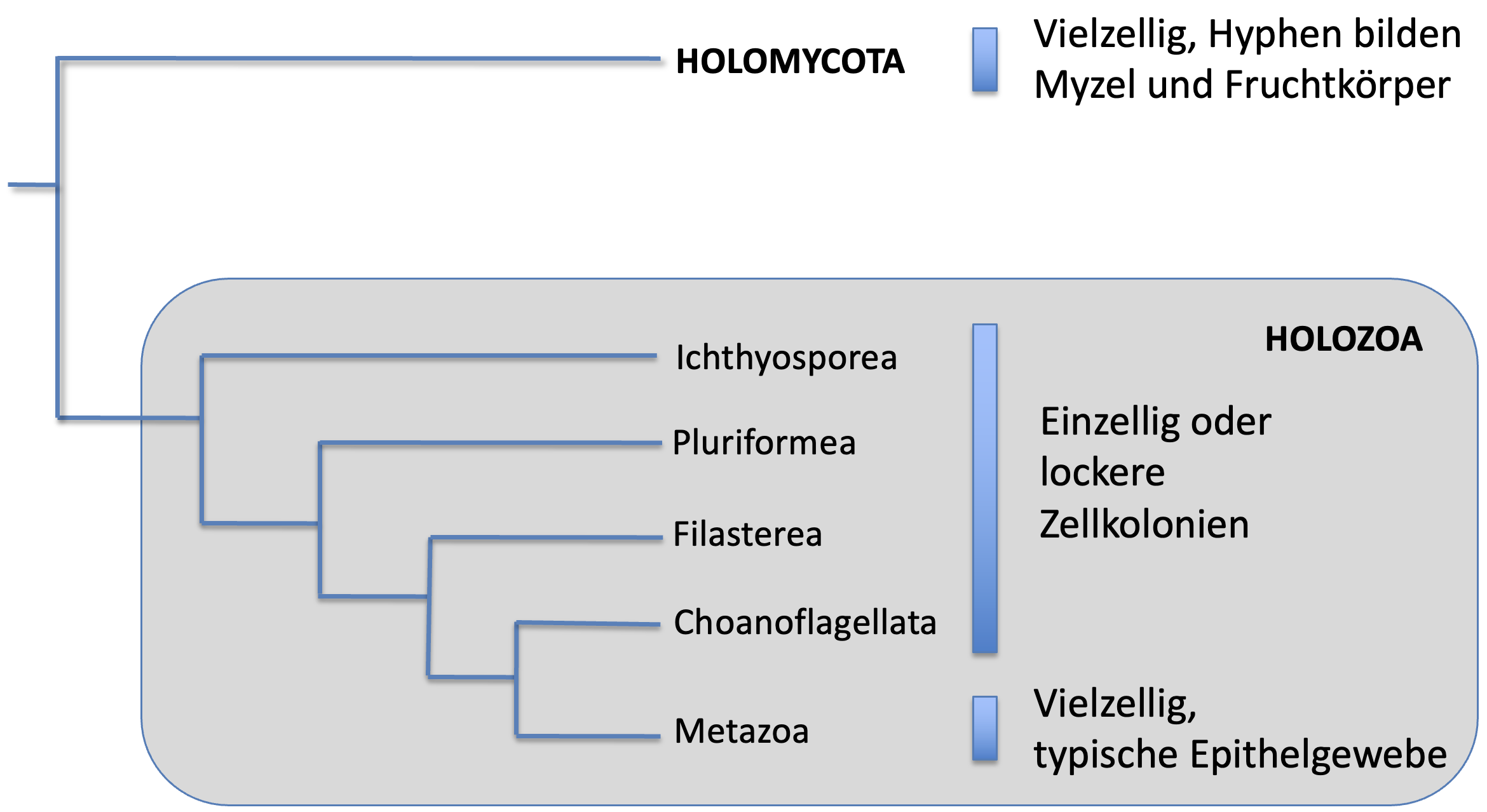
Die nächsten Verwandten der Metazoa innerhalb der Holozoa sind die Choanoflagellata (Kragengeißler) (siehe phylogenetischer Stammbaum in Abb. 1). Die Choanoflagellata sind heterotrophe, einzellige oder koloniale Flagellaten. Ihr deutscher Name deutet schon auf das Erkennungsmerkmal ihrer Zellen hin: den Kragen. Dabei handelt es sich um einen Ring aus Mikrovilli (fingerartige Zellauswüchse), der um eine zentrale Geißel herumsteht. Mit dieser Geißel können die Choanoflagellata einen Nahrungswasserstrom erzeugen, mit dem die Zellen Nahrungspartikel aus dem Wasser filtrieren können. Zellen mit einem nahezu identischen Aufbau, sog. Kragengeißelzellen oder Choanocyten, gibt es bei den ursprünglichsten Metazoa, den Schwämmen (Porifera). Die Schwämme sind echte Vielzeller, die neben den Kragengeißelzellen noch viele weitere Zelltypen besitzen und damit in ihrer Komplexität weit über die komplexesten Kolonien der Choanoflagellata hinausgehen. Trotzdem kann man davon ausgehen, dass Kolonien von Choanoflagellaten-ähnlichen Organismen als Ausgangspunkt der Evolution der Schwämme und der übrigen Metazoa angesehen werden können.
Ein weiterer Stamm ursprünglicher Metazoa sind die
Nesseltiere (Cnidaria). Bislang wurden etwa 15.000 Arten
wissenschaftlich beschrieben, die im Meer und im Süßwasser
vorkommen. Die Apomorphie der Nesseltiere sind die
Nesselzellen (Cnidocyten), die Kapseln (Cnidocysten)
enthalten, mit denen Nesselfäden und hochwirksame Gifte
explosiv freigesetzt werden können, und die der Verteidigung
und dem Beutefang dienen.
Neben den Porifera und Cnidaria gibt es drei weitere
ursprüngliche Metazoa-Stämme, nämlich die Placozoa, die
Ctenophora und die Monoblastozoa. Sie sind derzeit noch nicht
ausreichend erforscht, weswegen ihre Stellung im Stammbaum
noch unklar ist (incertae sedis). Die Scheibentiere (Placozoa)
sind zwar schon seit dem 19. Jahrhundert bekannt, aber erst
seit wenigen Jahren werden sie intensiver erforscht und es
konnten auch einige neue Arten entdeckt werden. Die
Rippenquallen (Ctenophora) wurden früher mit den Nesseltieren
als "Hohltiere" ("Coelenterata") zusammengefasst, bis man die
großen Unterschiede zwischen diesen beiden Gruppen besser
erkannte. Die Ctenophora sind offenbar eine sehr alte
Tiergruppe, deren Abstammungslinie sich schon sehr früh von
den anderen Metazoa getrennt hat, denn sie besitzen eine ganze
Reihe einzigartiger Merkmale, was sie zu echten "Rätseltieren"
macht. Andererseits wurde in den letzten Jahren vor
allem durch entwicklungsgenetische Forschungen klar, dass sie
neben den primären Keimblättern auch ein umfangreiches
Mesoderm entwickeln, und zwar sowohl Ektomesoderm als auch
Entomesoderm, und somit dieses charakteristische Merkmal der
Metazoa in typischer Ausprägung zeigen. Die Monoblastozoa
schließlich beinhalten nur eine einzige Art, Salinella salve,
die im 19. Jahrhundert aus einem Salzwerk (Saline) in
Südamerika beschrieben wurde, seither aber nicht wieder
gefunden werden konnte, trotz mehrerer Nachsuchen in der Nähe
des Originalfundorts.
Allen ursprünglichen Metazoa-Stämmen ist gemeinsam, dass sie
noch keine Körpersymmetrie aufweisen, bei der man definierte
linke und rechte Körperhälften festlegen könnte: es fehlt die
sog. bilaterale Symmetrie des Körpers. Bei den Porifera sind
die meisten Arten unsymmetrisch, und die meisten Cnidaria
haben runde oder zylindrische Körper
(Radiärsymmetrie), bei denen es nicht möglich ist, eine
linke oder rechte Körperhälfte zu definieren. Es gibt zwar bei
manchen Korallenarten schon Anzeichen einer bilateralen
Symmetrie, aber erst bei der Metazoa-Gruppe der Bilateria, zu
der auch der Mensch gehört, hat sich die bilaterale Symmetrie
(Bilateralsymmetrie) endgültig durchgesetzt. Man kann die
Bilateralsymmetrie sehr leicht bei sich selbst feststellen: es
gibt nur eine einzige Symmetrieachse, die den Körper in zwei
(mehr oder weniger) gleiche Seiten/Hälften teilt; das ist eine
echte Bilateralsymmetrie (lat. "bi-" = zwei, lat. "latus" =
Körperseite).
Zu den Bilateria gehören alle übrigen Stämme der Metazoa. Bei
den Bilateria sind nun alle Keimblätter und alle Gewebetypen
unumstritten vorhanden, und sie besitzen zahlreiche, für
bestimmte Aufgaben spezialisierte Organe bzw. Organsysteme.
Die Bilateria werden meist in zwei große Äste gegliedert, die
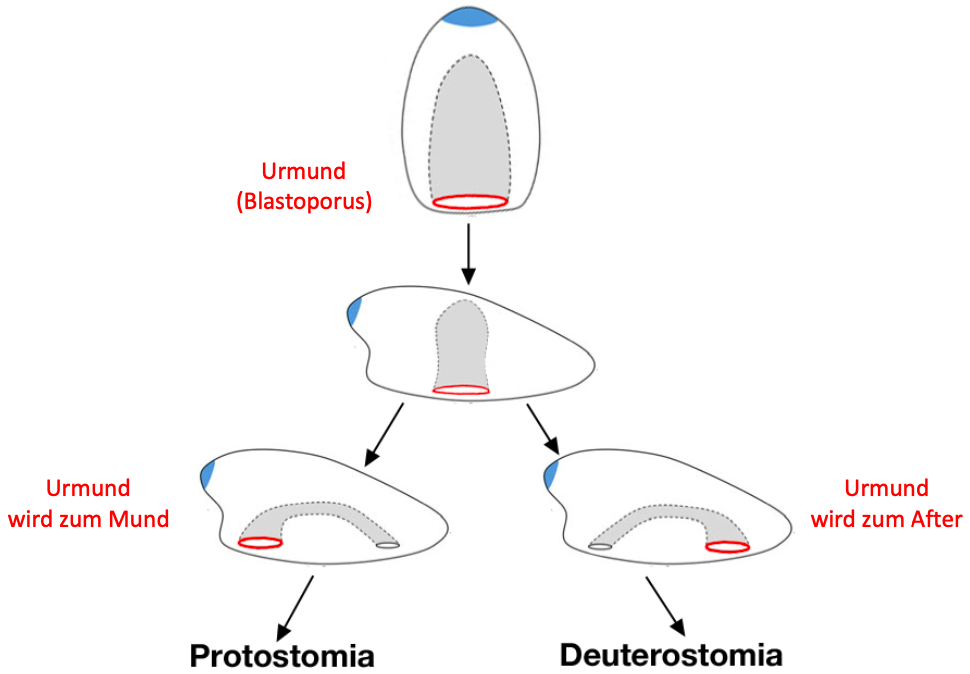
als Protostomia und Deuterostomia bezeichnet werden. Der Name
der zwei Großgruppen verrät bereits, was für den
entscheidenden Unterschied zwischen ihnen gehalten wird. Bei
den Protostomia (griech. protos = "zuerst, ursprünglich",
stoma = "Mund") entwickelt sich in der Embryonalentwicklung
der Blastoporus (Urmund) zum Mund und der After muss deshalb
als zusätzliche Öffnung neu gebildet werden. Die Protostomia
werden im Deutschen deshalb auch als Urmundtiere bezeichnet.
Bei den Deuterostomia ist es genau andersherum: der
Blastoporus wird zum After, so dass der Mund als zusätzliche
Öffnung neu gebildet werden muss; die Deuterostomia werden
deshalb auch Neumundtiere genannt (griech. deuteros =
"zweiter", stoma = "Mund"). Diese Vorstellung ist in Abb. 2
dargestellt und erklärt.
Allerdings hat sich inzwischen herausgestellt, dass diese
Vorstellung nicht widerspruchsfrei zutrifft. Zum einen
besitzen z. B. die Plattwürmer nur eine einzige Öffnung ihres
Verdauungssystems. Diese Öffnung ist also im Grunde noch der
Urmund, so dass die Plattwürmer nach diesem Merkmal weder
Proto- noch Deuterostomia sind. Alle Verwandtschaftsanalysen
belegen aber zweifelsfrei, dass sie zu den Protostomia
gehören. Zahlreiche neuere Studien haben außerdem gezeigt,
dass viele Protostomia ihren Darmtrakt während der
Embryonalentwicklung ganz anders entwickeln als nach der
Vorstellung der "Protostomie" vorhergesagt; sehr viele bilden
sowohl Mund als auch After aus dem Urmund (sog. Amphistomie)
und einige Arten bilden Mund und After sogar genauso wie es
typische Deuterstomia tun.
Es sind deshalb Zweifel an der phylogenetischen Aussagekraft
dieses Merkmals aufgekommen: weder die protostome Entwicklung
mit der Beibehaltung des Blastoporus als Mund, noch die
deuterostome Entwicklung bei der der Blastoporus zum After
wird und deshalb ein neuer Mund durchbrechen muss, scheinen
monophyletische Gruppen zu definieren. Was ist dann der
entscheidende Unterschied zwischen Proto- und Deuterostomia?
Oder müssen wir die Aufteilung der Bilateria in Protostomia
und Deuterostomia sogar aufgeben? Die Monophylie der
Protostomia kann jedenfalls auch durch andere Merkmale sehr
gut begründet werden, so dass an der Monophylie der
Protostomia derzeit keine Zweifel bestehen. Lediglich der Name
Protostomia ist dann nicht mehr passend, wird aber trotzdem
beibehalten, weil der Name allgemein im Gebrauch und sehr
bekannt ist und eine Umbenennung zu Verwirrung führen
würde.
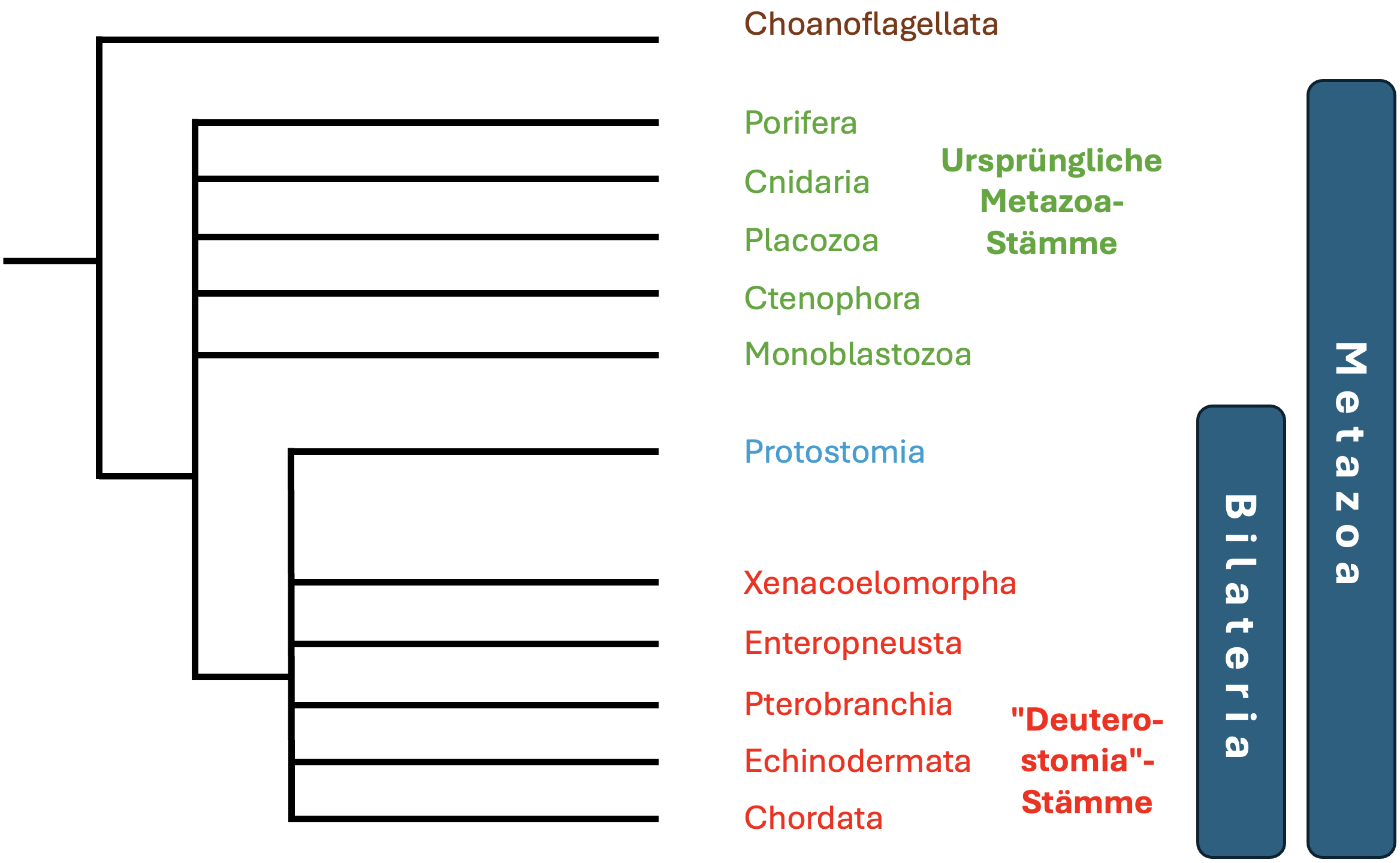
Die Begründung der Monophylie der Deuterostomia kann sich
jedoch kaum auf weitere Merkmale stützen. Es werden derzeit
vor allem zwei Merkmale diskutiert, die Apomorphien der
Deuterostomia darstellen könnten: (1) das mesodermale
Endoskelett und (2) der Kiemendarm. Die Homologie beider
Merkmale bleibt jedoch momentan unsicher.
Auch erste sehr genaue molekulare Analysen der
phylogenetischen Verwandtschaftsbeziehungen innerhalb der
Bilateria, deuten darauf hin, dass die Deuterostomia keine
monophyletische Gruppe sind (Philippe et al. 2019, Kapli et
al. 2021, Juravel et al. 2023). In Abb. 3 sind die
Deuterostomia deshalb nicht als monophyletische Gruppe
dargestellt; stattdessen sind die einzelnen Stämme separat
aufgeführt und die Verwandtschaftverhältnisse unaufgelöst
dargestellt.
"Deuterostomia", denen der typische Stützstab im Rücken (die
Rückensaite oder Chorda dorsalis) fehlt, und die deshalb als
ursprünglich angesehen werden, sind die Scheinplattwürmer
(Xenacoelomorpha), die Eichelwürmer (Enteropneusta), die
Flügelkiemer (Pterobranchia) und die Stachelhäuter
(Echinodermata). Die artenreichste Gruppe der "Deuterostomia"
sind die Rückensaitentiere (Chordata), zu denen auch der
Mensch gehört.
Die monophyletische Gruppe der Protostomia, besteht ihrerseits
aus zwei kleineren monophyletischen Gruppen: aus den Spiralia
und den Ecdysozoa (Abb. 4). Zu jeder der beiden Untergruppen
gehört eine Anzahl von Tierstämmen (Phyla). Die Spiralia
bestehen aus 16 Tierstämmen mit zusammen über 200.000
beschriebenen Arten. Namensgebende Apomorphie der Spiralia ist
eine besondere Form der Zellteilung während der frühen
Embryonalentwicklung, die sog. Spiralfurchung. Während dieser
frühen Furchungsteilungen wechselt die Stellung der
Teilungsspindel bei jeder Teilung, so dass die Furchungszellen
(Blastomeren) zum Beginn der Entwicklung spiralförmig
angeordnet sind. Im weiteren Verlauf der Entwicklung entsteht
eine bewimperte Schwimmlarve, die sich mittels Cilienbändern
fortbewegen kann. Dieser Larventypus, die sog.
Trochophoralarve, ist ebenfalls eine Apomorphie der Spiralia,
ist aber bei vielen Arten, vor allem natürlich bei
landlebenden Arten, vollständig zurückgebildet worden.
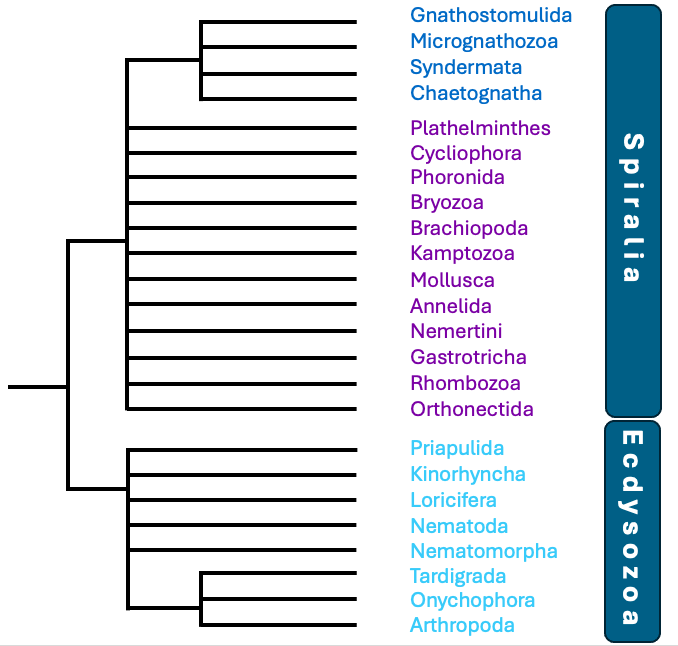
Die Ecdysozoa sind die artenreichste Tiergruppe: die acht
Tierstämme der Ecdysozoa zählen zusammen etwa 1,5 Millionen
beschriebene Arten, stellen damit fast 90% aller rezent
bekannten Tierarten und haben alle Lebensräume der Erde
erobert. Der mit Abstand artenreichste Stamm innerhalb der
Ecdysozoa sind die Arthropoda (Gliederfüßer, weit über 1
Million Arten). Neben den Arthropoda gehören noch die Nematoda
(Fadenwürmer, über 15.000 Arten), Nematomorpha (Saitenwürmer,
etwa 350 Arten), Priapulida (Priapswürmer, etwa 20 Arten),
Loricifera (Korsetttierchen, etwa 50 Arten), Kinorhyncha
(Hakenrüssler, etwa 200 Arten), Tardigrada (Bärtierchen, etwa
1.000 Arten) und Onychophora (Stummelfüßer, etwa 200 Arten) zu
den Ecdysozoa.
Die Ecdysozoa zeigen ursprünglich einen segmentierten Körper,
ein Merkmal, das die Ecdysozoa jedoch mit den Annelida und den
Chordata gemeinsam haben und das somit eine Plesiomorphie
darstellt. Nematoda, Nematomorpha, Priapulida und Loricifera
haben einen vereinfachten Körperbauplan ohne deutliche
Körpersegmentierung, was früher als ursprünglich interpretiert
wurde, heute aber als sekundäre Vereinfachung des ursprünglich
segmentierten Bauplans gilt. Dem ursprünglichen
Ecdysozoa-Bauplan am nächsten kommen stattdessen sehr
wahrscheinlich die Kinorhyncha. Sie zeigen eine deutliche
Segmentierung, die sich äußerlich durch die segmentalen
Cuticulaplatten des Exoskeletts zeigt und auch im Inneren
durch segmentale Muskelstränge und eine Bauchganglienkette
deutlich wird. Bei den Tardigrada kommen zu den segmentalen
Cuticulaplatten, Muskeln und Ganglien dann noch segmentale
Gliedmaßen (Laufbeine mit Krallen) hinzu. Einen ähnlichen
Bauplan wie die Tardigrada zeigen die Onychophora, bei denen
jedoch die Cuticulaplatten und die Segmentierung der
Muskulatur offenbar verlorengegangen sind und wieder ein
nahezu einheitlicher Hautmuskelschlauch vorliegt.
Die namensgebende Apomorphie der Ecdysozoa ist die Ecdysis,
die im deutschen meist als "Häutung" bezeichnet wird
("Häutungstiere"), obwohl bei der Ecdysis ausschließlich das
extrazelluläre Außenskelett (Exoskelett) ersetzt wird und die
eigentliche Haut nicht erneuert wird. Das Exoskelett, die
Cuticula, besteht aus Proteinen, die von den Epidermiszellen
in diskreten Schichten ausgeschieden werden. Jede Schicht
erhält durch Einlagerung zusätzlicher Komponenten in die
Protein-Matrix ganz spezielle Eigenschaften (Abb. 5):
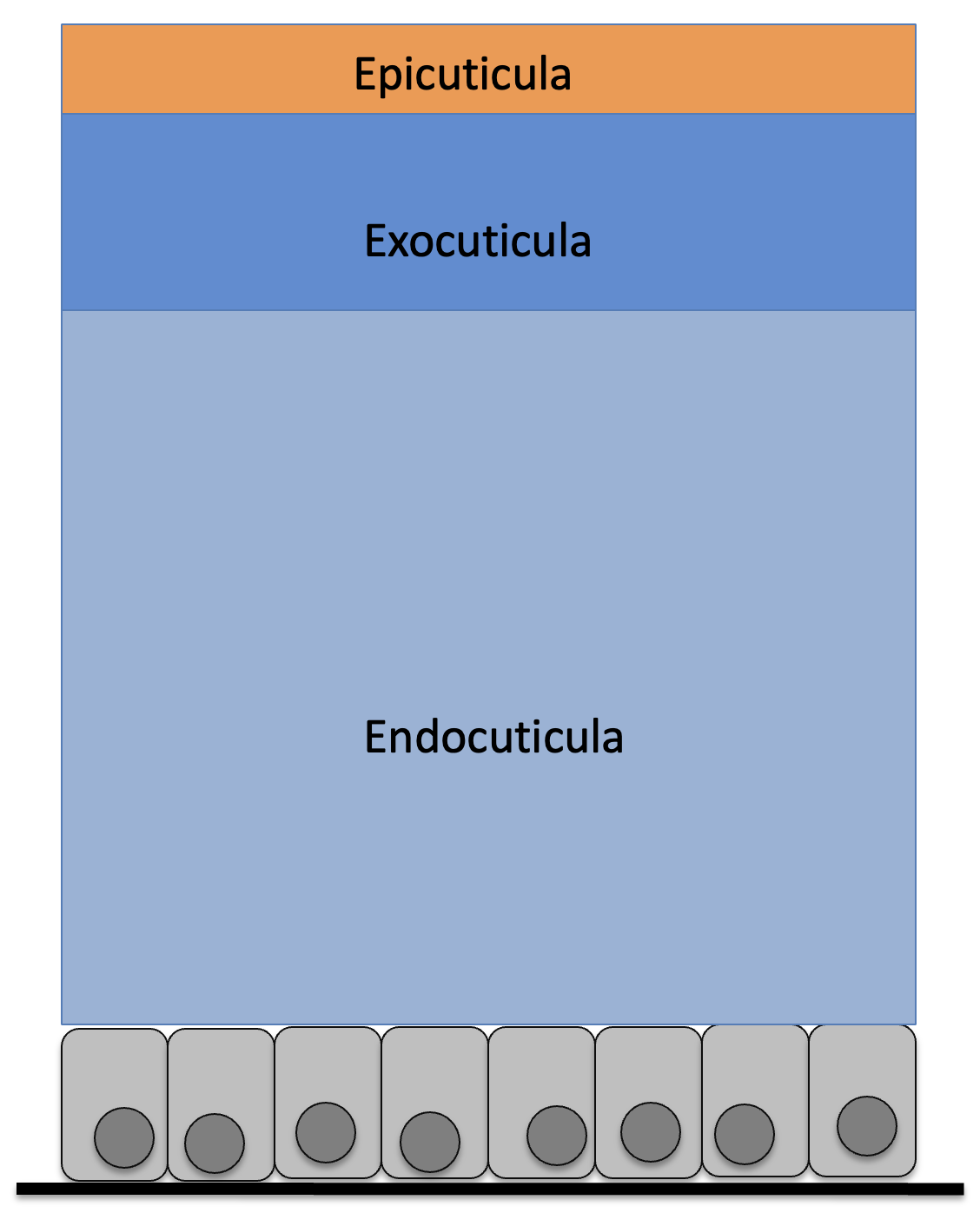
a) Sklerotisierung: die Proteine werden durch Chinone
miteinander vernetzt. Die Proteine bilden dadurch ein kaum
mehr bewegliches Netzwerk. Die betreffende Cuticulaschicht
wird starr und auch für Enzyme weitgehend unangreifbar (kann
also kaum enzymatisch abgebaut werden).
b) Bildung von Mikrofibrillen: die Proteine werden mit
filamentösen Polymeren (meist Chitin) verbunden; dadurch
entstehen zusammengesetzte fädige Strukturen (Mikrofibrillen)
aus Proteinen und Chitin. Die entsprechende Cuticulaschicht
erhält dadurch eine hohe Zug- und Druckfestigkeit, bleibt aber
(je nach Richtung der Fibrillen) elastisch.
c) Härtung: in das Netzwerk aus Proteinen werden Kalk (z. B.
beim Panzer vieler Krebstiere) oder Metalle (z. B. Kiefer
mancher Insekten) eingelagert. Die entsprechende
Cuticulaschicht erhält vorwiegend die Eigenschaften der zur
Härtung eingelagerten Komponente. Durch Härtung kann z. B.
eine Sklerotisierung weiter unterstützt werden (Bildung von
Körperpanzerplatten).
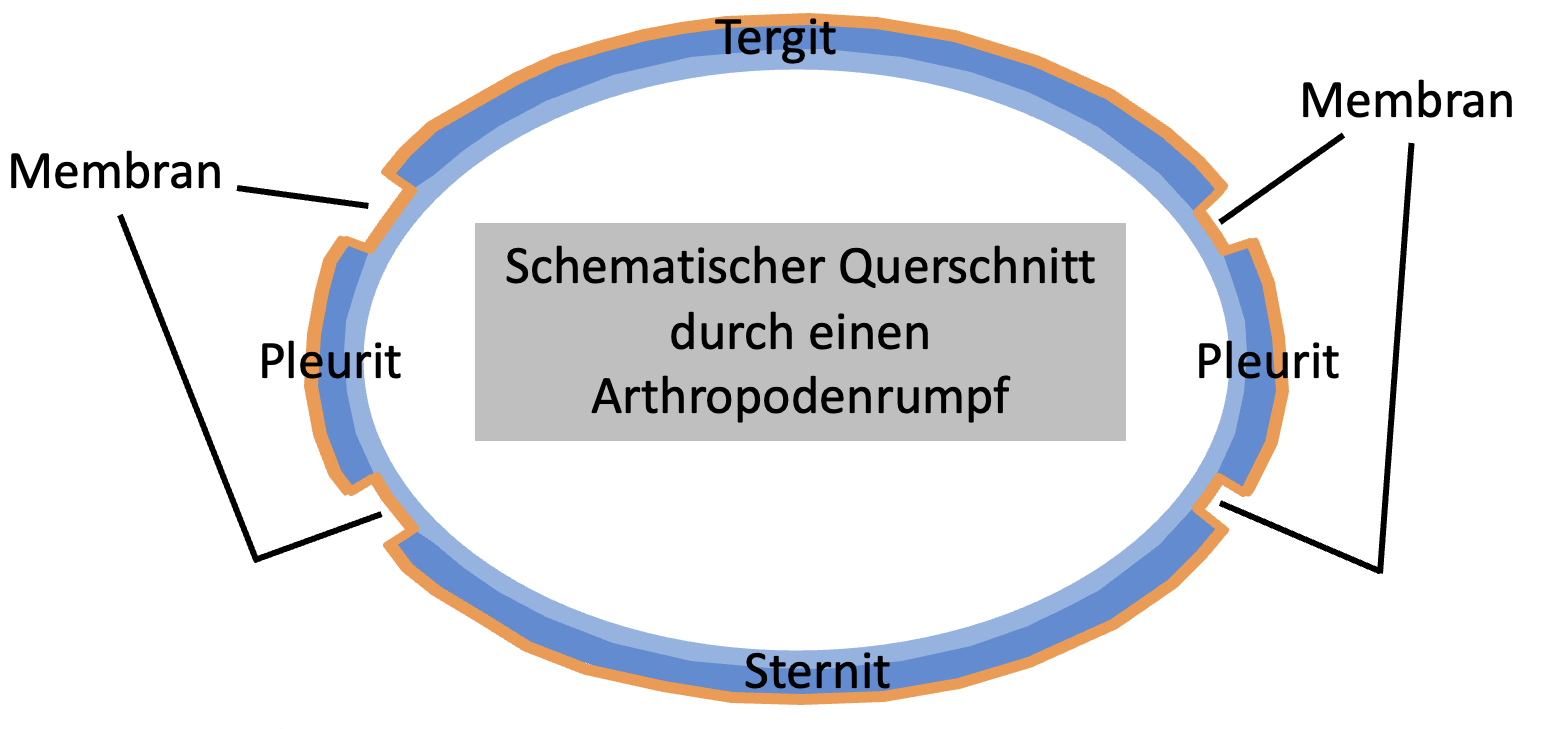
Die sehr dünne Epicuticula umgibt hauptsächlich als
Verdunstungsschutz den ganzen Körper und auch die elastische
Endocuticula ist um den ganzen Körper herum ausgebildet. Die
sklerotisierte Exocuticula ist jedoch nicht überall
ausgebildet, sondern nur dort wo eine besondere Verfestigung
der Cuticula nötig ist (z. B. als Panzerplatten zum Schutz,
sogenannte Sklerite ist. Cuticulabereiche ohne Exocuticula
sind elastisch und werden als Membranen bezeichnet (Abb. 6).
Bei den Ecdysozoa ist die sekundäre Leibeshöhle (Cölom)
nicht segmentiert. Bei vielen Ecdysozoa-Arten werden
Cölombläschen zwar embryonal angelegt, lösen sich dann aber im
weiteren Verlauf der Embryonalentwicklung auf, wodurch die
primäre Leibeshöhle und die sekundären Leibeshöhlen zu einer
einheitlichen Leibeshöhle, dem sogenannten Mixocöl,
verschmelzen. Bei den Arthropoda sind diese Verhältnisse klar
nachgewiesen. Bei den anderen Ecdysozoa-Stämmen ist die
Entwicklung des Mesoderms und damit auch die Entwicklung und
das Schicksal von sekundären Leibeshöhlen noch unzureichend
untersucht und die Entwicklung des Mixocöls noch nicht
zweifelsfrei nachgewiesen. In der Literatur wird deshalb oft
unbestimmt von einem "Pseudocöl" gesprochen.
Durch die Verschmelzung von primären und sekundären
Leibeshöhlen geht auch der Großteil der Cölomepithelien
verloren, wodurch kein vollständig geschlossenes
Kreislaufsystem gebildet werden kann; dorsal werden ein
Dorsalgefäß ("Herz") und wenige Gefäße angelegt. Ecdysozoa
haben deshalb ein überwiegend offenes Kreislaufsystem, in dem
sich also eine Mischung aus Cölomflüssigkeit und Blut
befindet, die als Hämolymphe bezeichnet wird. Die Hämolymphe
vereint dann die Funktionen der beiden Körperflüssigkeiten:
a) Stofftransport (Nährstoffe, Metabolite, Abbauprodukte,
Signalstoffe)
b) Immunabwehr
c) Wundverschluss
d) Hydraulik und Druck
e) Temperaturregelung (Wärmevermittler, Kühlflüssigkeit)
f) Atemgastransport
Juravel K, Porras L, Höhna S, Pisani D, Wörheide G. 2023. Exploring genome gene content and morphological analysis to test recalcitrant nodes in the animal phylogeny. PLoS One 18, e0282444.
Kapli P, Natsidis P, Leite DJ, Fursman M, Jeffrie N, Rahman IA, Philippe H, Copley RR, Telford MJ. 2021. Lack of support for Deuterostomia prompts reinterpretation of the first Bilateria. Science Advances 7, eabe2741.
Philippe H, Poustka AJ, Chiodin M, Hoff KJ, Dessimoz C, Tomiczek B, Schiffer PH, Müller S, Domman D, Horn M, Kuhl H, Timmermann B, Satoh N, Hikosaka-Katayama T, Nakano H, Rowe ML, Elphick MR, Thomas-Chollier M, Hankeln T, Mertes F, Wallberg A, Rast JP, Copley RR, Martinez P, Telford MJ. 2019. Mitigating anticipated effects of systematic errors supports sister-group relationship between Xenacoelomorpha and Ambulacraria. Current Biology 29, 1818-1826.e6.